
| Chap. I. | A General View of the Organiztion of the Vertebrata | |
| The Vertebrate Skeleton | 7 | |
| Chap. II. | The Muscles and the Viscera–A General View of the Organization of the Vertebrata | 44 |
| Chap. III. | The Provinces of the Vertebrata–The Class Pisces | 100 |
| Chap. IV. | The Class Amphibia | 149 |
| Chap. V. | The Classification and the Osteology of the Reptilia | 167 |
| Chap. VI. | The Classification and the Osteology of Birds | 233 |
| Chap. VII. | The Muscles and the Viscera of the Sauropsida | 256 |
| Chap. VIII. | The Classification and Organization of the Mammalia | 273 |
PREFACE.
[3] The present work is intended to provide students of comparative anatomy with a condensed statement of the most important facts relating to the structure of vertebrated animals, which have hitherto been ascertained. Except in a very few cases, I have intentionally abstained from burdening the text with references; and, therefore, the reader, while he is justly entitled to hold me responsible for any errors he may detect, will do well to give me no credit for what may seem original, unless his knowledge is sufficient to render him a competent judge on that head.
About two-thirds of the illustrations are original, the rest1 are copied from figures given by Agassiz, Bischoff, Burmeister, Busch, Carus, Dugès, Flower, Gegenbaur, Hyrtl, Von Meyer, Müller, Pander and D'Alton, Parker, Quatrefages, and Traquair.
A considerable portion of the book has been in type for some years; and this circumstance must be my excuse for appearing to ignore the views of several valued contemporaries. I refer more especially to those contained in recently-published works of Professors Flower and Gegenbaur.
London, September, 1881.
1 Namely, Figures 1, 6, 10, 11, 12, 13, 15, 18, 23, 26, 28, 29, 30, 31, 36, 39, 41, 42, 46, 50, 51, 54, 55, 56, 57, 58, 59, 60, 61, 75, 79, 82, 101, 107, 108, 109, 110.
[7]
A General View of the Organization of the Vertebrata–The Vertebrate Skeleton.
The Distinctive Characters of the Vertebrata.–The Vertebrata are distinguished from all other animals by the circumstance that a transverse and vertical section of the body exhibits two cavities, completely separated from one another by a partition. The dorsal cavity contains the cerebro-spinal nervous system; the ventral, the alimentary canal, the heart, and, usually, a double chain of ganglia, which passes under the name of the "sympathetic." It is probable that this sympathetic nervous system represents, wholly or partially, the principal nervous system of the Annulosa and Mollusca. And, in any case, the central parts of the cerebro-spinal nervous system, viz., the brain and the spinal cord, would appear to be unrepresented among invertebrated animals. For these structures are the results of the metamorphosis of a part of the primitive epidermic covering of the germ, and only acquire their ultimate position, in the interior of the dorsal tube, by the development and union of outgrowths of the blastoderm, which are not formed in the Invertebrata.1
Again, in the partition between the cerebro-spinal and vis[8]ceral tubes, certain structures, which are not represented in invertebrated animals, are contained. During the embryonic condition of all vertebrates, the centre of the partition is occupied by an elongated, cellular, cylindroidal mass–the notochord, or chorda dorsalis. And this structure persists throughout life in some Vertebrata ; but, in most it is more or less completely replaced by a jointed, partly fibrous and cartilaginous, and partly bony, vertebral column .
In all Vertebrata, that part of the wall of the visceral tube which lies at the sides of, and immediately behind, the mouth, exhibits, at a certain stage of embryonic development, a series of thickenings, parallel with one another and transverse to the axis of the body, which may be five or more in number, and are termed the visceral arches. The intervals between these arches become clefts, which place the pharyngeal cavity, temporarily or permanently, in communication with the exterior. Nothing corresponding with these arches and clefts is known in the Invertebrata.
A vertebrated animal may be devoid of articulated limbs, and it never possesses more than two pairs. These are always provided with an internal skeleton, to which the muscles moving the limbs are attached. The limbs of invertebrated animals are commonly more numerous, and their skeleton is always external.
When invertebrated animals are provided with masticatory organs, the latter are either hard productions of the alimentary mucous membrane, or are modified limbs. Vertebrated animals also commonly possess hard productions of the alimentary mucous membrane in the form of teeth, but their jaws are always parts of the walls of the parietes of the head, and have nothing to do with limbs.
All vertebrated animals have a complete vascular system. In the thorax and abdomen, in place of a single peri-visceral cavity in communication with the vascular system, and serving as a blood-sinus, there are one or more serous sacs. These invest the principal viscera, and may or may not communicate with the exterior–recalling, in the latter case, the atrial cavities of Mollusca.
In all Vertebrata, except Amphioxus, there is a single valvular heart, and all possess an hepatic portal system; the blood of the alimentary canal never being wholly returned directly to the heart by the ordinary veins, but being more or less completely collected into a trunk–the portal vein, which ramifies through and supplies the liver.
[9] The Development of the Vertebrata.–The ova of Vertebrata have the same primary composition as those of other animals, consisting of a germinal vesicle, containing one or many germinal spots, and included within a vitellus upon the amount of which the very variable size of the vertebrate ovum chiefly depends. The vitellus is surrounded by a vitelline membrane, and this may receive additional investments in the form of layers of albumen, and of an outer, coriaceous, or calcified shell.
The spermatozoa are always actively mobile, and, save some rare and exceptional cases, are developed in distinct individuals from those which produce ova.
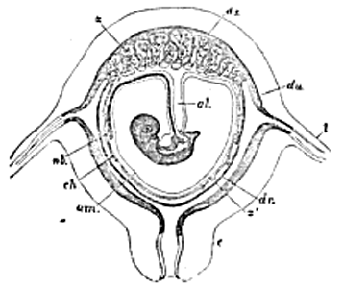
u, uterus; l, Fallopian tube; c, neck of the uterus; du, uterine decidua; ds , decidua serotina ; dr, decidua reflexa ; z, z' ; villi; am, amnion; nb , umbilical vesicle; al , allantois.
Impregnation may take place, either subsequently to the extrusion of the egg, when, of course, the whole development of the young goes on outside the body of the oviparous parent; or it may occur before the extrusion of the egg. In the latter case, the development of the egg in the interior of the body may go no further than the formation of a patch of primary tissue; as in birds, where the so-called cicatricula, or "tread," which is observable in the new-laid egg, is of this nature. Or, the development of the young may be completed [10] while the egg remains in the interior of the body of the parent, but quite free and unconnected with it; as in those vertebrates which are termed ovoviviparous. Or, the young may receive nourishment from its viviparous parent, before birth, by the close apposition of certain vascular appendages of its body to the walls of the cavity in which it undergoes its development.
The vascular appendages in question constitute the chief part of what is called the placenta, and may be developed from the umbilical vesicle (as in Mustelus among Sharks), or from the allantois and chorion (as in most mammals). At birth, they may be either simply detached from the substance of the parental organism, or a part of the latter may be thrown off along with them and replaced by a new growth. In the highest vertebrates, the dependence of the young upon the parent for nutrition does not cease even at birth; but certain cutaneous glands secrete a fluid called milk, upon which the young is fed for a longer or shorter time.
When development takes place outside the body, it may be independent of parental aid, as in ordinary fishes; but, among some reptiles and in most birds, the parent supplies the amount of heat, in excess of the ordinary temperature of the air, which is required, from its own body, by the process of incubation.
The first step in the development of the embryo is the division of the vitelline substance into cleavage-masses, of which there are at first two, then four, then eight, and so on. The germinal vesicle is no longer seen, but each cleavage-mass contains a nucleus. The cleavage-masses eventually become very small, and are called embryo-cells, as the body of the embryo is built up out of them. The process of yelk-division may be either complete or partial. In the former case, it, from the first, affects the whole yelk; in the latter, it commences in part of the yelk, and gradually extends to the rest. The blastoderm, or embryogenic tissue in which it results, very early exhibits two distinguishable strata–an inner, the so-called mucous stratum (hypoblast ), which gives rise to the epithelium of the alimentary tract; and an outer, the serous stratum (epiblast ), from which the epidermis and the cerebro-spinal nervous centres are evolved. Between these appears the intermediate stratum (mesoblast ),which gives rise to all the structures (save the brain and spinal marrow) which, in the adult, are included between the epidermis [11] of the integument and the epithelium of the alimentary tract and its appendages.
A linear depression, the primitive groove (Fig. 2, A, c ), makes its appearance on the surface of the blastoderm, and
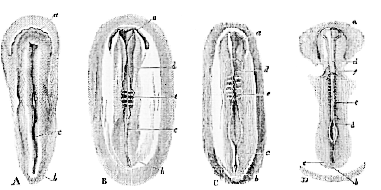
A, the first rudiment of the embryo; a its cephalic, b its caudal, end; c, the primitive groove.–B, the embryo further advanced; a, b, c, as before; d, the dorsal laminæ as yet developed only in the cephalic region, and not quite united in the middle line; e, the protovertebræ.–C, the letters as before. The dorsal laminæ have united throughout the greater part of the cephalic region, and are beginning to unite in the anterior spinal region.–D, embryo further advanced (the second day of incubation), the dorsal laminæ having united throughout nearly their whole length. The protovertebræ have increased in number, and the omphalomeseraic veins, f, are visible. The embryos are drawn of the same absolute length, but, in nature, the older are larger than the younger.
[12] the substance of the mesoblast along each side of this groove grows up, carrying with it the superjacent epiblast. Thus are produced the two dorsal laminæ, the free edges of which arch over toward one another, and eventually unite, so as to convert the primitive groove into the cerebro-spinal canal. The portion of the epiblast which lines this, cut off from the rest, becomes thickened, and takes on the structure of the brain, or Encephalon, in the region of the head; and of the spinal cord, or Myelon, in the region of the spine. The rest of the epiblast is converted into the epidermis.
The part of the blastoderm which lies external to the dorsal laminæ forms the ventral laminæ ; and these bend downward and inward, at a short distance on either side of the dorsal tube, to become the walls of a ventral, or visceral, tube. The ventral laminæ carry the epiblast on their outer surfaces, and the hypoblast on their inner surfaces, and thus, in most cases, tend to constrict off the central from the peripheral portions of the blastoderm. The latter, extending over the yelk, encloses it in a kind of bag. This bag is the first-formed and the most constant of the temporary, or fœtal, appendages of the young vertebrate, the umbilical vesicle.
While these changes are occurring, the mesoblast splits, throughout the regions of the thorax and abdomen, from its ventral margin, nearly up to the notochord (which has been developed, in the mean while, by histological differentiation of the axial indifferent tissue, immediately under the floor of the primitive groove), into two lamellæ . One of these, the visceral lamella, remains closely adherent to the hypoblast, forming with it the splanchnopleure, and eventually becomes the proper wall of the enteric canal; while the other, the parietal lamella , follows the epiblast, forming with it the somatopleure, which is converted into the parietes of the thorax and abdomen. The point of the middle line of the abdomen at which the somatopleures eventually unite, is the umbilicus.
The walls of the cavity formed by the splitting of the ventral laminæ acquire an epithelial lining, and become the great pleuroperitoneal serous membranes.
The Fœtal Appendages of the Vertebrata.–At its outer margin, that part of the somatopleure which is to be converted into the thoracic and abdominal wall of the embryo, grows up anteriorly, posteriorly, and laterally, over the body of the embryo. The free margins of this fold gradually approach one another, and, ultimately uniting, the inner layer of the fold becomes converted into a sac filled with a clear [13] fluid, the Amnion; while the outer layer either disappears or coalesces with the vitelline membrane, to form the Chorion (Fig. 3).
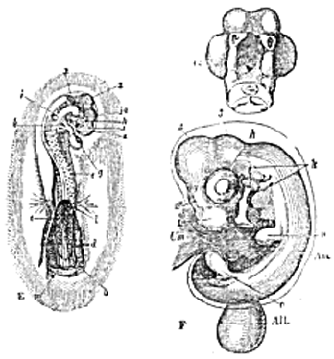
Fig. 3.–Later stages of the development of the body of a Fowl than those represented in Fig. 2.–E, embryo at the third day of incubation, g, heart, h, eye; i, ear; k, visceral arches and clefts; l, m, anterior and posterior folds of the amnion which have not yet united over the body; 1, 2, 3, first, second, and third cerebral vesicles; 1a, vesicle of the third ventricle.–F, embryo of the fifth day of incubation. The letters as before, except n, o, rudiments of the anterior and posterior extremities; Am, amnion; All (the allantois, hanging down from its pedicle); Um, umbilical vesicle.–G, under-view of the head of the foregoing, the first visceral arch being cut away.
Thus the amnion encloses the body of the embryo, but not the umbilical sac. At most, as the constricted neck, which unites the umbilical sac with the cavity of the future intestine, becomes narrowed and elongated into the vitelline duct , and as the sac itself diminishes in relative size, the amnion, increasing in absolute and relative dimensions, and becoming distended with fluid, is reflected over it (Fig. 1).
A third fœtal appendage, the Allantois, commences a single or double outgrowth from the under surface of the meso[14]blast, behind the alimentary tract; but soon takes the form of a vesicle, and receives the ducts of the primordial kidneys, or Wolffian bodies. It is supplied with blood by two arteries, called hypogastric, which spring from the aorta; and it varies very much in its development. It may become so large as to invest all the rest of the embryo, in the respiratory, or nutritive, processes of which it then takes an important share.
The splitting of the ventral laminæ, and the formation of a pleuroperitoneal cavity, appear to take place in all Vertebrata. Visually, there is a more or less distinct umbilical sac; but in fishes and Amphibia there is no amnion; and the allantois, if it is developed at all, remains very small in these two groups.
Reptiles, birds, and mammals have all these fœtal appendages. At birth, or when the egg is hatched, the amnion bursts and is thrown off, and so much of the allantois as lies outside the walls of the body is similarly exuviated; but that part of it which is situated within the body is very generally converted, behind and below, into the urinary bladder, and, in front and above, into a ligamentous cord, the urachus, which connects the bladder with the front wall of the abdomen. The umbilical vesicle may either be cast off, or taken into the interior of the body and gradually absorbed.
The majority of the visceral clefts of fishes and of many Amphibia remain open throughout life; and the visceral arches of all fishes (except Amphioxus), and of all Amphibia, throw out filamentous or lamellar processes, which receive branches from the aortic arches, and, as branchiæ , subserve respiration. In other Vertebrata all the visceral clefts become closed and, with the frequent exception of the first, obliterated; and no branchiæ are developed upon any of the visceral arches.
In all vertebrated animals, a system of relatively or absolutely hard parts affords protection, or support, to the softer tissues of the body. These, according as they are situated upon the surface of the body, or are deeper seated, are called exoskeleton, or endoskeleton .
The Vertebrate Endoskeleton.–This consists of connective tissue, to which cartilage and bone may be added in various proportions; together with the tissue of the notochord and its sheath, which cannot be classed under either of these heads. The endoskeleton is distinguishable into two independant portions–the one axial, or belonging to the head and trunk; the other, appendicula , to the limbs.
The axial endoskeleton usually consists of two systems of [15] skeletal parts, the spinal system, and the cranial system, the distinction between which arises in the following way in the higher Vertebrata :
The primitive groove is, at first, a simple straight depression, of equal diameter throughout; but, as its sides rise and the dorsal laminæ gradually close over (this process commencing in the anterior moiety of their length, in the future cephalic region), the one part becomes wider than the other, and indicates the cephalic region (Fig. 4, A). The notochord, which underlies the groove, terminates in a point at a little distance behind the anterior end of the cephalic enlargement, and indeed under the median of three dilatations which it presents. So much of the floor of the enlargement as lies in front of the end of the notochord, bends down at right angles to the rest; so that the anterior enlargement, or anterior cerebral vesicle , as it is now called, lies in front of the end of the notochord; the median enlargement, or the middle cerebral vesicle , above its extremity; and the hinder enlargement, or the posterior cerebral vesicle, behind that extremity (Fig. 4, D and E). The under surface of the anterior vesicle lies in a kind of pit, in front of, and rather below, the apex of the notochord, and the pituitary gland is developed in connection with it. From the opposite upper surface of the same vesicle the pineal gland is evolved, and the part of the anterior cerebral vesicle in connection with which these remarkable bodies arise, is the future third ventricle.
Behind, the posterior cerebral vesicle passes into the primitively tubular spinal cord (Fig. 4, A). Where it does so, the head ends, and the spinal column begins; but no line of demarcation is at first visible between these two, the indifferent tissues which ensheath the notochord passing without interruption from one region to the other, and retaining the same character throughout.
The first essential differentiation between the skull and the vertebral column is effected by the appearance of the protovertebræ . At regular intervals, commencing at the anterior part of the cervical region, and gradually extending backward, the indifferent tissue on each side of the notochord undergoes a histological change, and gives rise to more opaque, quadrate masses, on opposite sides of the notochord (Fig. 2, B, C). Each pair of these gradually unite above and below that structure, and send arched prolongations into the walls of the spinal canal, so as to constitute a protovertebra.
No protovertebræ appear in the floor of the skull, so that, [16] even in this early stage, a clear distinction is drawn between the skull and the spinal column.
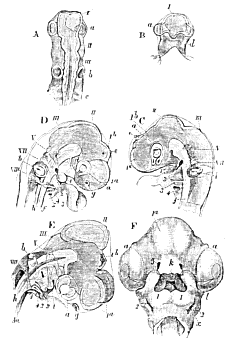
I, II, III, first, second, and third cerebral vesicles; Ia, vesicle of the cerebral hemisphere; Ib vesicle of the third ventricle; a, rudiments of the eyes and optic nerves; b, of the ears; g, of the olfactory organs; d, the infundibulum; e, the pineal gland; c, protovertebræ; h, notochord; 1, 2, 3, 4, 5, visceral arches; V, VII, VIII, the trigeminal, portio dura, and ninth and tenth pairs of cranial nerves; k, the nasal process; l, the maxillary process; x, the first visceral cleft. A, B, upper and under views of the head of a Chick at the end of the second day. C, side-view at the third day. D, side-view at seventy-five hours. E, side-view of the head of a Chick at the fifth day, which has been subjected to slight pressure. F, head of a Chick at the sixth day, viewed from below.
1 It is possible that an exception to this rule may be found in the Ascidians. The tails of the larvæ of these animals exhibit an axial structure, which has a certain resemblance to a vertebrate notochord; and the walls of the pharynx are perforated much as in Amphioxus.
[169] The Reptilia.–This class is divisible, by well-defined characters, into the following groups:
A. The dorsal vertebræ (which, like all the other vertebræ, are devoid of transverse processes) are not movable upon one another, nor are the ribs movable upon the vertebræ (Pleurospondylia). Most of the dorsal vertebræ and ribs are restrained from motion by the union of superficial long plates, into which they pass, to form a carapace.
Dermal bones, usually nine in number, one of which is median and symmetrical, and the others lateral and paired, are developed in the ventral walls of the thorax and abdomen, forming a plastron.
I.–Chelonia.
B. The dorsal vertebræ (which have either complete, or rudimentary, transverse processes) are movable upon one another, and the ribs upon them. There is no plastron.
a. The dorsal vertebræ have transverse processes, which are either entire, or only very imperfectly divided into terminal facets (Erpetospondylia).
a. The transverse processes are long; the limbs well developed, with the digits united by the integument into a paddle; the sternum and sternal ribs are absent or rudimentary.
II.–Pleisosauria.
b. The transverse processes are short, and sometimes rudimentary; the limbs present or absent; when they are fully developed, the digits are free, and there is a well- developed sternum with sternal ribs.
[170] a. A pectoral arch and urinary bladder.
III.–Lacertilia.
b. No pectoral arch, and no urinary bladder.
IV.–Ophidia.
b. The dorsal vertebræ have double tubercles in the place of transverse processes (Perospondylia.) The limbs are paddle-like.
V.–Ichthyosauria.
c. The anterior dorsal vertebræ have elongated and divided transverse processes, the tubercular being longer than the capitular division (Suchospondylia.)
a. With only two vertebræ in the sacrum.
VI.–Crocodilia.
b. With more than two vertebræ in the sacrum.
a. The manus without a prolonged ulnar digit
1. The hind-limb saurian.
VII.–Dicyodontia.
2. The hind-limb ornithic.
VIII.–Ornithoscelida.
b. The manus having an extremely prolonged ulnar digit.
IX.–Pterosauria.
I shall describe the exoskeletal, endoskeletal, and dental systems of the chief groups of the Reptilia, under the several heads here enumerated, and I shall then give an account of these systems in Aves. But modifications in the myology, neurology, splanchology, and development of the two classes may be conveniently considered together, under those several heads, in another chapter.
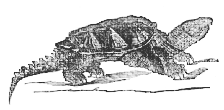
1. The Chelonia.–The Tortoises and Turtles are those reptiles which most nearly approach the Amphibia, though they depart very widely not merely from the amphibians but from the ordinary vertebrate type, in some respects.
A horny epidermic exoskeleton is absent in the soft Tortoises (Trionyx), the bodies of which are covered by a soft integument; but, in the other Chelonia, the epidermis is modified into horny plates, which constitute the so-called "Tortoise-shell," and have in general a very definite arrangement. The dorsal surface of the body presents three series of central plates, of which five are in the middle and four upon each side (4, 5, 4). The margins of the dorsal shield are guarded by twenty-four or twenty-five plates; one on the middle line in fronts called nuchal; one or two behind, pygal; and eleven [171] on each side, marginal. The ventral shield sometimes presents an anterior median scale; but, more usually, there are six pairs, disposed symmetrically. It will be seen, presently, that these epidermic plates by no means correspond with the bony dermal ossifications. In addition to these principal plates, smaller scale-like patches of horny epidermis are developed on other parts of the body, and on the limbs.
The dermal ossifications may best be described in connection with the endoskeleton.
The presacral vertebræ are few in number. In the Green Turtle (Chelone midas) there are eight cervical, and ten dorsal, in front of the sacrum, which is composed of two vertebræ. In all the cervical vertebræ the neurocentral sutures persist; there are no transverse processes, or ribs, and the spines are low or obsolete. The first vertebra, or atlas, is a ringlike bone, composed of three pieces, one basal and two superolateral. The second is a true axis vertebra, the central part of the centrum of the atlas ossifying apart, as an odontoid bone, and attaching itself to the front face of the centrum of the second vertebra.
The other cervical vertebræ are remarkable for the singular variety in the disposition of their articular cups and bells.
Thus the third is opisthocœlous; the fourth, biconvex; the fifth, procœlous; the sixth, also procœlous, but the posterior face is nearly flat, and very broad; in the seventh, both the anterior and the posterior faces are very broad and flattened, the posterior being the more convex. The eighth cervical vertebra is procœlous, and differs from the rest by the expansion of its neural spine, and by the arching backward of its postzygapophyses over the convex prezygapophyses of [172] the first dorsal vertebra, upon which the former play backward and forward.
All the cervical vertebræ are very freely movable upon one another, and confer great flexibility on the neck. In striking contrast with this arrangement, the ten following vertebræ have flattened faces, firmly united by cartilage. If any one of these vertebræ, from the second to the ninth, be examined, it will be found that the elongated centrum is only loosely united with the neural arch, and that the summit of the neural arch is continuous with a broad flat plate of bone, which forms one of the eight median elements of the carapace, or neural plates (Fig. 62, V).
There are no transverse processes, but a rib is articulated between the centrum and the neural arch. At a short distance from its attachment, this rib passes into a broad plate of bone, which extends upward to unite suturally with the neural plate; and, in front and behind, becomes similarly connected with preceding and succeeding costal plates, The rib may be traced along the under-surface of the costal plate, beyond the outer margin of which it protrudes; and its free extremity is received into a pit in an elongated prismatic dermal ossification, which forms one of a series of marginal plates (Fig. 62, M).
C1, centrum; V, expanded neural plate; C, costal plate; R, rib; M, marginal plate; P, lateral element of the plastron.
The first dorsal vertebra differs from the others in many respects. The anterior face of its centrum is concave, and looks downward and forward, while its prezygapophyses are much prolonged, in order to articulate with the convex posterior face of the centrum and, prolonged postzygapophyses of the last cervical vertebræ. The spinous process of this vertebra does not pass into the bony nuchal plate of the carapace, which lies above it (Fig. 63, Nu), and its rib does not expand into a costal plate, but becomes connected with the costal plate of the second dorsal vertebra. The neural arch of this [173] vertebra is shorter, from before backward, than its centrum; and the neural arch of the second dorsal vertebra extends forward and overlaps the centrum of the first, for the space thus left unoccupied. The rib of the second vertebra is also carried forward, and articulates not only with its own centrum and neural arch, but with the posterior edge of the centrum of the first vertebra.
These arrangements are repeated by the other dorsal vertebræ and ribs, up to the ninth inclusive; but, in the tenth, the neural arch occupies only the anterior half of the centrum of its own vertebra, and the rib is very small, and has no costal plate.
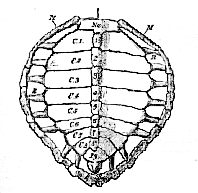
Nu, neural plate, M, marginal plates; R, ribs; 1-8, neural plates; C.1-C.8, costal plates; Py, pygal plates
The union of the neural and costal plates of the eight dorsal vertebræ, from the second to the ninth inclusively, gives rise to the principal part of the carapace, or dorsal moiety of the bony shell of the Chelonian. The first and the tenth dorsal vertebræ contribute nothing to the carapace, their small ribs merely becoming attached to the costal plates behind and before them.
In front of the first neural plate, and joined with it by a serrated suture, lies a large nuchal plate (Fig. 63. Nu), which [174] forms the anterior median boundary of the carapace. This neural plate sends down from its under-surface a median process, which is joined by ligament with the expanded neural spine of the eighth cervical vertebra. Behind the eighth neural plate, three other median pygal plates (Fig. 63, Py) succeed one another. The anterior two of these are united by sutures with the eighth neural and costals, and with one another; but the third is connected externally only with the marginal plates. All three are perfectly distinct from the subjacent vertebræ.
The sides of the carapace are completed, between the nuchal and pygal plates, by eleven marginal plates (Fig. 63, M) on each side. Eight of these receive the ends of the ribs of the second to the ninth dorsal vertebræ in the way already described.
There is no doubt that the neural, the pygal, and the marginal plates of the carapace are membrane-bones, developed in the integuments quite independently of either the vertebra or the ribs. But it appears that the neural plates and the costal plates exist, as expansions of the cartilages of the neural spines and ribs of the primitive vertebræ, before ossification takes place. This being the case, the neural and costal plates are vertebral and not dermal elements, however similar they may seem to be to the nuchal, pygal, and marginal plates. But this ultimate similarity of bones of totally distinct origin is not more remarkable here than in the case of the skull where the parietal and frontal bones stand in the same relation to the supra-occipital bone as the neural and pygal plates do to the neural plates of the carapace.
There are no sternal ribs, and no trace of a true sternum has yet been discovered in the Chelonia. The plastron is wholly composed of membrane-bones, which are developed in the integument, and lie, in part, in front of, and, in part, behind, the umbilicus of the fœtus. The latter, at least, therefore, belong to the abdomen, and the plastron is a thoracico-abdominal structure.
In the turtle the plastron consists of nine pieces–one median and anterior, four lateral and paired (Fig. 64). These pieces may be named–the median, entoplastron; the first later, epiplastron; the second, hyoplastron; the third, hypoplastron; and the fourth, xiphiplastron.1 The entoplastron [175] and the two epiplastra correspond with the median and lateral thoracic plates of the Labyrinthodont Amphibia, and very probably answer to the interclavicle and clavicles of other Vertebrata.
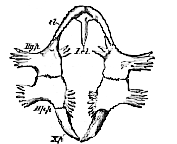
I.cl., interclavicle; cl, clavicle; Hy.p., hyoplastron; Hp.p., hypoplastron; Xp., xiphiplastron.
The sacrum consists of two vertebræ. The expanded sacral ribs are not anchylosed with the centra and arches of their vertebræ.
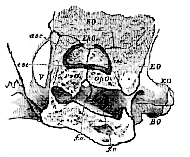
f.o., fenestra ovalis; f.p., fenestra rotunda; esc, asc, psc, external, anterior, and posterior semicircular canals.
The tail is flexible, and consists of procœlous vertebræ. [176] The anterior caudal vertebræ have no transverse processes, but possess ribs which may not become anchylosed with the centra. Thus the tail and the neck are the only regions of the spinal column of a Chelonian which are flexible.
In the skulls of the Chelonia all the bones, except the mandible and the hyoidean arch, are immovably united together.
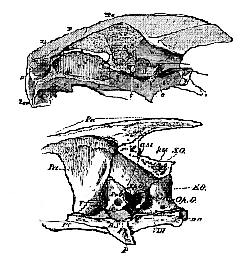
The upper figure represents the entire skull with the outline of the brain in situ; the lower gives a larger view of the inner face of the bones of the posterior moiety of the skull.
In the occipital segment of the adult, the supra-occipital is united with the epiotic, but the ex-occipital usually remains perfectly distinct from the opisthotic. The basisphenoid is large and distinct. The alisphenoidal region remains unossified; but the large parietals send down a prolongation on each side, which plays the part of an alisphenoid. Neither the presphenoid nor the orbitosphenoids are represented by [177] bone, but there are large frontals. In the periotic capsule the large proötic and the opisthotic (Cuvier's occipitate externe) remain distinct bones, but the epiotic unites with the supra-occipital.
The naso-ethmoidal cartilage largely persists; but it by comes covered above and at the sides by a large bone, which meets with its fellow in the middle link, and occupies the position of the lachrymal, prefrontal, and nasal. The premaxillæ are small, and usually united together. There is a single vomer, produced downward into a median internasal plate, which expands below, and joins the palatine plate of the palatine bone.
Above the posterior and upper part of the orbit lies a post-frontal, and, behind this, a squamosal is phased at the sides of the periotic capsule, and above the large quadrate bone. The post-frontal and squamosal occupy the upper part of the temporal region of the skull. Below these, a quadrato-jugal and a jugal connect the quadrate bone with the large maxilla.
In some genera, as Chelone and Chelydra, the skull possesses a sort of false roof, formed by the expansion of a median ridge, developed from the parietal bones, into a broad plate, which becomes suturally united with the post-frontals and squamosals.
The quadrate bone is firmly fixed to the sides of the periotic region of the skull, and ends below in a strong condyle for the mandibles. The long and broad pterygoid bones unite with one another in the middle line, and are firmly fixed to the base of the skull, as in Plesiosauria and Crocodilia. They unite only with the upper part of the quadrate bone, as in the latter reptiles.
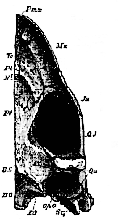
The palatines are firmly united with the pterygoids, behind, and with the vomer above and in front. They are prolonged downward, and develop a short palantine plate, which unites with the produced and expanded lower edge of the vomer, to bound the posterior nares. (Fig. 67, Vo, N 1).
[178] The dentary pieces of the two rami of the mandible are represented by one bone, as in Birds.
The hyoidean apparatus consists of a broad plate of cartilage with two longer anterior, and two shorter posterior, ossified cornua. The cornua have no direct connection with the skull.
The pectoral and pelvic arches appear, at first sight, to have a very anomalous position in the Chelonia, inasmuch as they seem to be situated inside and not outside, the skeleton of the trunk. But since the plastron does not answer to the sternum of other Vertebrata, but to part of the dermal skeleton, the anomaly does not really exist on the ventral side. And, as to the dorsal side, the pectoral and pelvic arches of the fœtal Chelonian are at first situated in front of, or behind, and external to, the ribs, as in other Vertebrata. It is only as development advances, that the first costal plate extends over the scapula, and the hinder costal plates over the ilium.
The pectoral arch is ossified in such a manner that the scapula and precoracoid form one bone, while the coracoid remains distinct. The free ends of the coracoid and precoracoid are usually connected together by a fibro-cartilaginous band, representing the epicoracoidal cartilage in Lacertilia. There is no clavicle, unless the epiplastra and entoplastron represent that bone.
The carpus of the Chelonia contains nine primary ossicles, as in the Urodela–three in the proximal row, one central, and five distal–and these almost always remain distinct.
There are five digits, the members of the phalanges of which present no constancy.
The pelvis contains the usual bones. The pubes (which are very large) and the ischia meet respectively in a long symphysis; and, sometimes, the foramina obturatoria are completed, internally, by the meeting of the bony pubes and ischium of each side in the middle line.
The pelvis is not usually united with either the carapace or the plastron, but in Chelys, Chelonia, and some other genera, the ilia unite by synchondrosis, or anchylosis, with the last costal plate, and the pubis and ischium with the xiphisternal plates, so that the pelvis becomes firmly fixed between the carapace and plastron.
The proximal row of the tarsal bones consists usually of an astrogalus, formed by the union of the tibiale and intermedium, and of a fibulare or calcaneum. In Chelydra, there is a [179] centrale. In Chelone, Emys, Testudo, and Trionyx the centrale is united with the astragalus; and in Emys, the calcaneum coalesces into the astragalus, so that the proximal portion of the tarsus consists of one bone. In the distal series the two fibular tarsals are united into a cuboid bone. There are five digits, and the fifth metatarsal has a peculiar form, as if bent upon itself at right angles, in the middle of its length.
The Chelonia are divisible into the Testudinea, the Emydea, the Trionychoidea, and the Euereta.
1. The Testudinea have the horny jaws naked and cutting, or denticulated. The eyes are lateral, the tympanic membrane is exposed, the short and thick limbs have the toes (all of which have nails) bound together by the integument. The horny plates of the carapace and plastron are well developed.
The Land Tortoises belong to this division. The carapace is usually very convex, and sometimes (as in the genus Pyxis) the anterior part of the plastron is movable, and can be shut up like a lid. In Cinyxis, the hinder part of the carapace is similarly mobile.
2. The Emydea have usually, horny cutting jaws, uncovered by lips; the tympanum exposed, and the limbs more slender than in the Testudinea, With five-clawed digits, which are only united by a web. The horny plates of the carapace and plastron are well developed.
These are the River and Marsh Tortoises. They are further divisible into two groups, in the one of which, the Terrapenes, the pelvis is free, the neck bends in a vertical plane, and the head is almost completely hidden by the carapace when retracted (Emys, Cistudo, Chelydra). In Cistudo, Cinostenum, and Staurotypus, the hinder part of the plastron is mobile. In the other division, the Chelodines, the pelvis is fixed to the carapace and plastron, the neck bends sideways, and the head cannot be completely retracted under the carapace (Chelys, Chelodina.)
3. In the Trionychoidea (Mud or Soft Tortoises), the jaws have an external cutaneous lip; the nasal organ is prolonged into a kind of snout, and the head is covered by a soft skin without any visible tympanic membrane. The limbs are flattened, somewhat finlike, and pentadactyle; but only three digits have nails. The integument develops no horny plates, [180] but is quite soft. The costal plates are shorter than in other Chelonia, and the marginal ossicles are either rudimentary or absent.
The genera Gymnopus, Cryptopus, and Cycloderma, constitute this division; they all inhabit the fresh waters of hot latitudes.
The Euereta, or Turtles, have an exposed, hooked, horny beak, with a blunt snout. The tympanum is hidden by the integument. The limbs, of which the anterior pair are much the longer, are converted into paddles, the digits being much flattened and elongated, and immovably united together by the integument; only one or two nails are developed. The skin of the body is either rugose (Sphargis), or covered with thick epidermic plates (Chelone).
The two genera composing this group inhabit the seas of warmer climates.
The Chelonia are first known to occur, with certainty, in the Lias. The older forms are, in many respects, intermediate between the Euereta and the Trionychoidea, but present no approximation to any other order of Reptilia.
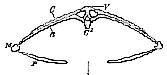
II. The Plesiosauria.–In some of the Plesiosauria, the head, not much than one-twelfth or one-thirteenth of the length of the body, is mounted upon a neck as longs in proportion, as that of a Swan; but in others, the head is large and massive and the neck much shorter. The hind-limbs are longer than the fore-limbs, and there is a comparatively short tail The integument was certainly devoid of any scutes; and was, probably, smooth and possessed no scales.
The cervical vertebræ may exceed forty in number, though they are generally fewer; and as none of the ribs appear to have been connected with the sternum, or if such connection existed it cannot now be traced, it becomes difficult to distinguish between cervical and dorsal vertebrae and one is obliged to have recourse to some method of separating the two, differing from that already adopted. Now, in these animals, the neurocentral suture persists for a considerable period, if not throughout life; and the surfaces for the articulation of the cervical ribs, which are at first altogether below the neurocentral sutures, gradually rise, in the posterior parts of the neck, until they first are cut by, and then rise above the suture. It is very convenient, and harmonizes very well with some facts to be mentioned by-and-by in the structure of the Crocodilia, to take the last of the vertebræ in which the [181] costal articular surface is cut by the neurocentral suture, as the last of the cervical series.
The two anterior cervical vertebræ, as thus defined, constitute the atlas and axis, and are frequently anchylosed together. The centre of the other cervical vertebræ have slightly concave anterior and posterior surfaces; well-developed neural arches; anterior and posterior oblique processes, or zygapophyses, of the ordinary character; and stout, but somewhat short, spinous processes. The centrum presents, upon each side, an oval rugose pit, sometimes more or less divided into two facets. This is the costal articular surface, which has been already adverted to. Into it fits the thickened head of a costal rib, which may have corresponding facets, but is otherwise undivided. The rib is continued backward into a short and straight body, and the angle, or the part at which the neck and the body of the rib join, is produced forward, so that the cervical ribs of the Plesiosauria have a strong general resemblance to those of the Crocodilia. In the posterior part of the neck and the anterior part of the dorsal region, the ribs become somewhat longer, and lose their anterior processes, gradually acquiring the rounded and curved form of ordinary ribs. Their proximal ends remain simple, and the facets, with which they articulate, become raised, and thrown outward, as transverse processes, developed from the arches of the vertebræ. (Fig. 68, C.)
In the anterior dorsal vertebræ these transverse processes rapidly acquire their full length; and they are continued under this form, descending somewhat lower upon the arches of the vertebræ toward the sacrum, to the end of the dorsal region. The neural spines acquire greater length, the zygapophyses are well developed, and the articular surfaces of the centra retain the form which they possessed in the cervical region. There are usually between twenty and twenty-five dorsal vertebræ. The sacral vertebræ are two, and resemble the others, except that the sacral ribs are large and broad for the attachment of the ilium. The caudal vertebræ, usually between thirty and forty in number, become, as usual, reduced to little more than centra at the end of the tail; but, in the fore part of the tail, they have well-developed spines and articular processes, with ribs which become anchylosed to the bodies of the vertebræ, only late in life. Well-developed chevron-bones are attached between the ventral margins of successive centra of the caudal vertebræ.
As has been mentioned there appear to be no sternal ribs, [182] but there is a well-developed system of ossification of the wall of the abdomen, arranged in transverse rows from before backward; each row consists of a median bone, slightly bent upon itself, thick in the middle, and think at each end–and of six other bones, three on each side, which are elongated and
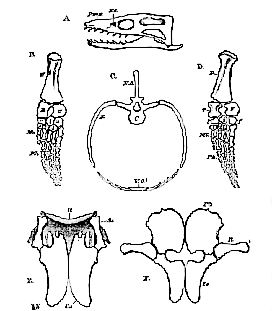
[183] pointed at each end, and so disposed that their pointed ends overlap one another. (Fig. 68, C.)
In some Plesiosauria, as already stated, the skull (Fig. 68, A) is very small in proportion to the body, not having more than a twelfth, or a thirteenth, of the length of the latter; but, in other species, the skull is much larger. The snout is tapering and depressed, and the nasal apertures are situated, not at its extremities, but just in front of the orbits–the latter, like the supra-temporal fossæ, being wide. The occipital condyle is almost wholly developed from the stout basi-occipital. The ex-occipitals give off elongated parotic processes, and the basisphenoid is a thick bone, which ends in front in a long rostrum.
There is a well-marked parietal foramen, and the parietals send off comparatively short processes backward, which become connected with the large squamosals. Tile latter unite with the post-frontals, which separate the orbits from the temporal fossa, and the orbit is completed behind by the junction of the post-frontal with the jugal. The jugal bone is continued backward into a slender bar, which extends as far back as the lower end of the quadrate, and probably contains a quadratojugal, so that there is a distinct infra-temporal fossa. The most obvious circumstance in which the skull of Plesiosaurus differs from that of most Reptilia is in the great size of the premaxillaries, which constitute a large proportion of the snout.
The undersurface of the skull is rarely well exposed in its anterior part; posteriorly, it exhibits a broad and long expansion, formed by the pterygoid bones, which unite in the middle line, and send processes outward and backward to the quadrate bone. On each side of the middle line of this region of the skull, is seen an ovoidal fossa or depression. The pterygoids are continued forward, and are united externally with transverse bones, and more anteriorly with flattened palatine bones. When the fore-part of the under-surface of the skull is exposed, two other fossæ are visible, one on each side of the middle line, bounded behind by the palatine bones, and separated by what appear to be the vomers. I conceive that these are the true posterior nares, and that the posterior apertures are simply spaces left between the pterygoid bones and the basis cranii.
At the sides of the base of the skull, specimens of Plesiosaurus occasionally exhibit two styliform bones, which lie parallel with the axis of the skull; these may be parts of the [184] hyoidean apparatus. No trace of any sclerotic ring has been found.
The teeth of the Plesiosaurus are sharp-pointed, curved, and the outer surfaces of their crowns striated. Each tooth is lodged in a distinct alveolus, with which, as in the Crocodilia, it does not become anchylosed.
The pectoral arch (Fig. 68, E) is one of the most remarkable parts of the organization of the Plesiosaurus. It consists, in the first place, of two very large coracoids, the long axes of which are parallel with one another, while their inner edges meet, without overlapping, throughout the greater part of their extent. In this respect they differ from any of the Lacertilia, which are provided with well-developed limbs. In these the long axes of the coracoids always cut one another at a large angle, open posteriorly–a circumstance which results from the manner in which the coracoids are received into grooves in the anterolateral edges of the rhomboidal part of the sternum. Hence it would appear that the Plesiosauria, like the Chelonia, did not possess any thing corresponding to this rhomboidal part of the sternum, but that the intercoracoid part of the sternum was either absent, or reduced to a mere band, as in some Batrachia.
The scapulæ are unlike the corresponding organs in any other reptile. The glenoidal end, stout and strong, is continued horizontally forward and inward, as a bony prism, with a somewhat concave inner edge and flat inferior surface. The outer surface, rising up at right angles to the ventral surface, gives rise to a well-defined edge; at a short distance from the glenoidal end, the part of the bone which bears this surface is produced upward and backward, into a low recurved plate. This part appears to represent the proper body of the scapula in other Reptiles, while the horizontal prolongation answers to that preglenoidal process of the scapula, which extends forward and inward as a free bony bar in many Lacertilia–for example, Iguana.
In well-preserved specimens, a broad hoop of substance (Fig. 68, E, a), which seems to have been but imperfectly ossified, extends across the middle line of the body, from the preglenoidal process of one scapula to that of the other, and is continued backward in the middle line, to the junction of the two coracoids. This corresponds very nearly in form and position to the epicoracoidal ossifications of the Lacertilia, combined with the clavicles and interclavicles; but I have never been able to detect any distinct clavicular, or intercla[185]vicular, elements in any Plesiosaurus, though they appear to have been well developed in Nothosaurus.
The humerus is a stout bone–prismatic, and with a rounded head at its proximal end, flattened and broad distally. (Fig. 68, B) Its anterior margin is nearly straight, or even slightly convex, while the posterior is concave. Distally, it presents two facets, meeting at an angle, with which the broad and short radius and ulna articulate. The ulna differs in shape from the radius, being convex posteriorly, and concave in front. The two bones are of equal length, and much shorter than the humerus. There are six rounded carpal bones,2 arranged in two rows; and to these succeed five digits, composed of metacarpals and phalanges, which are elongated and constricted in the middle. The middle digits have numerous phalanges.
The pelvic arch has very large dimensions, in correspondence with the size of the hind-limb, which is usually longer than the fore-limb. (Fig. 68, F and D.) The ilium is a vertically elongated bone, narrower below than above, where it becomes connected with the sacral ribs. Inferiorly, it unites with the pubis and with the ischium, to form the acetabulum. The pubes are very broad quadrate bones, much larger than the ischia, and they meet in a median symphysis. The ischia, triangular and expanded, also unite in a ventral symphysis. The femur resembles the humerus in its general form, although both its sides are straighter, and the other bones of the hind-limb are so like those of the fore-limb, as to need no special description.
There can be little doubt that all the bones of the limbs were, like those of the Cetacea, enclosed within a common sheath of integument, so as to form a paddle.
Such is the general organization of the skeleton of the Plesiosauria, which are long extinct animals, entirely confined to the Mesozoic Rocks, from the Trias to the Chalk, inclusive. They may be divided into two groups, according as they are Triassic, or Post-Triassic, in age.
The Post-Triassic group contains the genera Plesiosaurus and Pliosaurus, the different species of which appear to differ in little more than the proportions of the head to the trunk, and the relative length and decree of excavation of the centra [186] of the vertebræ. In the species which have been named Pliosaurus, the vertebræ are wide in proportion to their length, and deeply excavated in front and behind. Pliosaurus attained gigantic dimensions, paddles of some individuals reaching a length of not less than six feet.
The Triassic genera, Nothosaurus, Simosaurus, Pistosaurus (for a knowledge of the organization of which we are chiefly indebted to the labors of Hermann von Meyer), appear to have differed from Plesiosaurus principally in the following respects:
The connection of the neural arches with the centra of the vertebræ seems to have been looser. The supra-temporal fossæ in the skull appear to have been larger in proportion. In these animals, the under-surface of the skull has the same structure as in Plesiosaurus, but apparently lacks the posterior fossæ; while there is no doubt whatsoever that the true posterior nares are situated far forward, in the position assigned to them in Plesiosaurus.
The pectoral arch of Nothosaurus, again, presents a very interesting deviation from the Plesiosaurian type. The coracoids, indeed, are greatly expanded, and meet by their inner edges, so that the rhomboidal part of the sternum seems to have been wholly absent, and the scapulæ have a horizontal prolongation, not quite so long as in Plesiosaurus, with an upstanding proper scapular part of corresponding shape. But then the ends of these preglenoidal processes are connected together by, and indeed suturally united with, a stout, curved, transverse bar of bone, consisting of three pieces, one small and median, and two very large and lateral, all united firmly together by sutures. There can be little doubt that the constituents of this bony bar correspond with the interclavicles and clavicles of Lacertilia and Ichthyosauria.
[208] V. The Ichthyosauria.–In its general form Ichthyosaurus presents a good deal of resemblance to a Cetacean. The head is enormous, and passes at once into the trunk, so that there is no more appearance of a neck than in a Porpoise, and the body tapers off behind, much as would happen in the latter animal were it devoid of a caudal fin. Indeed, there is some reason to suspect that the tail of Ichthyosaurus may have been provided with a sort of fin-like expansion of the integument. This fish-like body was propelled, like that of the Plesiosaurus, by four paddles; but the anterior paddles were placed close behind the head, and were, generally, very much larger than the posterior ones.

[209] The spinal column is only distinguished into two regions, caudal and precaudal, inasmuch as the ribs, beginning at the anterior part of the neck, are continued, without being connected with the sternum, to the posterior end of the body; and there is no sacrum. The caudal region, however is dis-
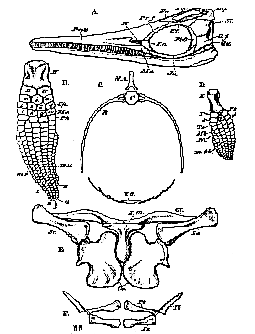
A, the head;
B, the fore-limb; H, humerus; R. radius; U, ulna; r, i. u, radiale, intermedium, ulnare; Cp, carpalia; 1, 2, 3, 4, 5, digits; m.r m.u radial and ulnar marginal ossicles.
C. a dorsal vertebra, with the ribs (R) and ventral ossifications (V.O).
D. the hind-limb; F, femur; T, tibia; Fb, fibula; t, i, f, tibiale, intermedium, fibulare; Ts, tarsalia; Mt, metatarsalia; Ph, phalanges; m, tb, tibial marginal ossicles.
E, the pectoral arch, seen from the ventral side;
F, the same aspect of the pelvic arch.
[210]tinguished by the chevron-bones which are attached beneath its vertebrae The vertebræ of Ichthyosauria in general have certain characters by which they differ from those of all other Vertebrata. (Fig. 76, C.) Not only are the centra flattened disks, very much broader and higher than they are long, and deeply biconcave (circumstances in which they resemble the vertebræ of some Labyrinthodonts and Fishes), but the only transverse processes they possess are tubercules, developed from the sides of these centra; and the neural arches are connected with two flat surfaces, one on each side of the middle line of the upper surface of the vertebræ by mere synchondroses. The neural arches themselves are forked bones, with only rudiments of zygapophyses, and in the greater part of the body do not become articulated with one another at all.
In the cervical region, if one may call "neck" the most anterior part of the vertebral column, the front part of the lateral surface of each vertebra presents two separate elevations, or articular surfaces, which are at first situated in the upper half of the lateral surface. Toward the posterior half of the dorsal region they descend, and, gradually approaching one another, coalesce into one in the caudal vertebræ. The form of the proximal ends of the ribs corresponds with the arrangement of these tubercles; for, where they are separate, the proximal end of the rib is forked. The lower fork, or capitulum, goes to the capitular, or lower, tubercle, and the upper branch, or tuberculum, to the upper, or tubercular, elevation. In the caudal region, where the articular surface is single, the proximal end of the rib is also undivided. In the caudal region the ribs are short and straight, but in the precaudal region they are stout and curved, and much longer in the middle than at either end of the series. The atlas and axis resemble the other vertebræ in their general form: but a wedge-shaped bone is, as it were, let in between their opposed lower edges; and a similar bone, attached to the underpart of the concave face of the centrum of the atlas, serves to complete the cup for the occipital condyle.
The skull of Ichthyosaurus (Fig. 76, A) is remarkable for the great elongation and tapering form of the snout, the huge orbits, the great supra-temporal fossæ, and the closing over of the infra-temporal fossæ by plates of bone. Again, the two rami of the mandible are united in a symphysis, which, for length, is comparable to that observed in the modern Gavials and in the ancient Teleosauria. The basi-occipial bone fur[211]nishes the round articular condyle to the first vertebra, and becomes very stout and thick in front. It appears to have been anchylosed neither with the basisphenoid nor with the basi-occipital. The latter bones are adapted to its sides, and, together with the supra-occipital, which is interposed between them above, circumscribe the occipital foramen. The basisphenoid, a deep and stout bone, is produced in front into a long and slender parasphenoidal rostrum. There do not appear to have been any ossified alisphenoids. The parietals remain separate throughout life; and, in some species, not merely present a great parietal foremen close to the coronal suture, but are completely divided by a median fissure. Ossified presphenoids and orbitosphenoids appear to have been altogether absent, and the frontal bones are relatively small. The proötic bones are, as usual, situated in front of the ex-occipitals, and between the latter and them there may sometimes be discerned a conical bone with a broad base, which appears to be fitted in between the exoccipital and the proötic. If this bone were not so large, it might well be regarded as a stapes, but it is possible that, as Cuvier suggests, it answers to the separate opisthotic of the Chelonia
In the naso-premaxillary segment, the nasal bones, continuing the direction of the frontals, attain considerale size, but the premaxillæ make up by far the greater part of the snout. The maxillæ are reduced, as in birds, to comparatively small and slender rod-like bones, bounding only a fraction of the gape. The vomers are elongated, and situated in the middle line on the underside of the snout.
The nostrils are small apertures close to the orbits, bounded by the nasal, lachrymal, and premaxillary bones.
On each side of the frontal there is a large prefrontal, which passes back above to meet the post-frontal, and thus bound the orbit. Below, the maxilla is connected with a jugal. From the post-frontal to the jugal, the posterior margin of the orbit is constituted by a distinct, curved, postorbital bone (Fig.76, A, Pt. O). A broad and flat quadratojugal (Q,j.) passes from the end of the jugal to the louver end of the quadrate, and covers in the lower and posterior part of the infra-temporal fossa. The space between this bone, the post-orbital, the post-frontal and the squamosal, is occupied by another flattened bone (Fig. 76, A, St.), which Cuvier calls the temporal, but which does not appear to have any precise homologue among other Reptilia.....
1 Believing the plastron to answer to the sternum of other Vertebrata, anatomists have termed these elements of the plastron entosternum, episternum, hyosternum, hyposternum, and xiphisternum. 2 It may be a question whether the fourth distal bone in the carpus and tarsus (Fig. 68, B and D) belongs to carpus and tarsus, or to metacarpus and metatarsus; or whether it is formed by the confluence of elements belonging to both regions.
|
THE
HUXLEY
FILE

|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
{kind=link}