An Introduction to the Classification of Animals (1866)
 An Introduction to the Classification of Animals (1869)
An Introduction to the Classification of Animals (1869)
An Introduction to the Classification of Animals (1869)
An Introduction to the Classification of Animals (1869)| Chapter 1 | On Classification in General | 1 |
| Chapter 2 | The Characters of the Classes of the Invertebrata | 6 |
| Chapter 3 | The Characters of the Classes of the Vertebrata | 59 |
| Chapter 4 | On the Arrangement of the Classes into Larger Groups | 75 |
| Chapter 5 | The Subclasses and Orders into which the Classes of the Vertebrata Are Divisible | 87 |
| Chapter 6 | The Orders into Which the Classes of the Invertebrata Are Divisible | 115 |
| Glossary | 13 |
[1]
By the classification of any series of objects, is meant the actual, or ideal, arrangement together of those which are like and the separation of those which are unlike; the purpose of this arrangement being to facilitate the operations of the mind in clearly conceiving and retaining in the memory, the characters, of the objects in question.
Thus, there may be as many classifications of any series of natural, or of other, bodies, as they have properties or relations to one another, or to other things; or, again, as there are modes in which they may be regarded by the mind: so that, with respect to such classification as we are here concerned with, it might be more proper to speak of a classification than of the classification of the animal kingdom.
The preparations in the galleries of the Museum of the Royal College of Surgeons are arranged upon the basis laid down by John Hunter, whose original collection was intended to illustrate the modifications which the great physiological apparatuses undergo in the animal series: the classification which he adopted is a classification by organs, and, as such, it is admirably adapted to the needs of the comparative physiologist.
But the student of the geographical distribution of animals, regarding animated creatures, not as diverse modifications of one [2] great physiological mechanism, but in relation to one another, to plants and to telluric conditions, would, with equal propriety, dispose of the contents of a Zoological Museum in a totally different manner; basing his classification, not upon organs, but on distributional assemblages. And the pure palæontologist, looking at life from yet another distinct point of view, would associate animal remains together on neither of these principles, but would group them according to the order of their succession in Time.
Again, that classification which I propose to discuss in the present pages, is different from all of these: it is meant to subserve the comprehension and recollection of the facts of animal structure; and, as such, it is based upon purely structural considerations, and may be designated a Morphological Classification. I shall have to consider animals, not as physiological apparatuses merely; not as related to other forms of life and to climatal conditions; not as successive tenants of the earth; but as fabrics, each of which is built upon a certain plan.
It is possible and conceivable that every animal should have been constructed upon a plan of its own, having no resemblance whatsoever to the plan of any other animal. For any reason we can discover to the contrary, that combination of natural forces which we term Life might have resulted from, or been manifested by, a series of infinitely diverse structures: nor, indeed, would anything in the nature of the case lead us to suspect a community of organization between animals, so different in habit and in appearance, as a porpoise and a gazelle, an eagle and a crocodile, or a butterfly and a lobster. Had animals been thus independently organized, each working out its life by a mechanism peculiar to itself, such a classification as that which is now under contemplation would obviously be impossible; a morphological, or structural, classification plainly implying morphological or structural resemblances in the things classified.
As a matter of fact, however, no such mutual independence of animal forms exists in nature. On the contrary, the different members of the animal kingdom, from the highest to the lowest, are marvellously connected. Every animal has a something in common with all its fellows: much, with many of them; [3] more, with a few; and, usually, so much with several, that it differs but little from them.
Now, a morphological classification is a statement of these gradations of likeness which are observable in animal structures, and its objects and uses are manifold. In the first place, it strives to throw our knowledge of the facts which underlie, and are the cause of, the similarities discerned into the fewest possible general propositions, subordinated to one another, according to their greater or less degree of generality; and in this way it answers the purpose of a memoria technica, without which the mind would be incompetent to grasp and retain the multifarious details of anatomical science.
But there is a second and even more important aspect of morphological classification. Every group in that classification is such in virtue of certain structural characters, which are not only common to the members of the group, but distinguish it from all others; and the statement of these constitutes the definition of the group.
Thus, among animals with vertebræ, the class Mammalia is definable as those which have two occipital condyles, with a well-ossified basi-occipital; which have each ramus of the mandible composed of a single piece of bone and articulated with the squamosal element of the skull; and which possess mammæ and non-nucleated red blood-corpuscles.
But this statement of the characters of the class Mammalia is something more than an arbitrary definition. It does not merely mean that naturalists agree to call such and such animals Mammalia: but it expresses, firstly, a generalization based upon, and constantly verified by, very wide experience; and, secondly, a belief arising out of that generalization. The generalization is that, in nature, the structures mentioned are always found associated together: the belief is, that they always have been, and always will be, found so associated. In other words, the definition of the class Mammalia is a statement of a law of correlation. or coexistence, of animal structures, from which the most important conclusions are deducible.
For example: if a fragmentary fossil be discovered, consisting of no more than a ramus of a mandible and that part of the [4] skull with which it articulated, a knowledge of this law may enable the palæontologist to affirm, with great confidence, that the animal of which it formed a part suckled its young and had non-nucleated red blood-corpuscles; and to predict that, should the back part of that skull be discovered, it will exhibit two occipital condyles and a well-ossified basi-occipital bone.
Deductions of this kind, such as that made by Cuvier in the famous case of the fossil opossum of Montmartre, have often been verified, and are well calculated to impress the vulgar imagination; so that they have taken rank as the triumphs of the anatomist. But it should carefully be borne in mind, that, like all merely empirical laws, which rest upon a comparatively narrow observational basis, the reasoning from them may at any time break down. If Cuvier, for example, had had to do with a fossil Thylacinus instead of a fossil Opossum, he would not have found the marsupial bones, though the inflected angle of the jaw would have been obvious enough. And so, though, practically, any one who met with a characteristically mammalian jaw would be justified in expecting to find the characteristically mammalian occiput associated with it; yet, he would be a bold man indeed, who should strictly assert the belief which is implied in this expectation, viz., that at no period of the world's history did animals exist which combined a mammalian occiput with a reptilian jaw, or vice versa.
Not that it is to be supposed that the correlations of structure expressed by these empirical laws are in any sense accidental, or other than links in the general chain of causes and effects. Doubtless there is some very good reason why the characteristic occiput of a Mammal should be found in association with mammal and non-nucleated blood-corpuscles; but it is one thing to admit the causal connection of these phenomena with one another, or with some third; and another thing to affirm that we have any knowledge of that causal connexion, or that physiological science, in its present state, furnishes us with any means of reasoning from the one to the other.
Cuvier, the more servile of whose imitators are fond of citing his mistaken doctrines as to the nature of the methods of palæontology against the conclusions of logic and of common [5] sense, has put this so strongly that I cannot refrain from quoting his words.1
"But I doubt if any one would have divined, if untaught by observation, that all ruminants have the foot cleft, and that they alone have it. I doubt if any one would have divined that there are frontal horns only in this class: that those among them which have sharp canines for the most part lack horns.
"However, since these relations are constant, they must have some sufficient cause; but since we are ignorant of it, we must make good the defect of the theory by means of observation: it enables us to establish empirical laws, which become almost as certain as rational laws, when they rest on sufficiently repeated observations; so that now, whoso sees merely the print of a cleft foot may conclude that the animal which left this impression ruminated, and this conclusion is as certain as any other in physics or morals. This footprint alone, then, yields to him who observes it, the form of the teeth, the form of the jaws, the form of the vertebræ, the form of all the bones of the legs, of the thighs, of the shoulders, and of the pelvis of the animal which has passed by: it is a surer mark than all those of Zadig."
1 "Ossemens fossiles," ed. 4me, tome lr, p. 184.
[13]
Multitudinous forms of sponges exist in both salt and fresh waters. Up to the last few years we were in the same case, with respect to this class, as with the Gregarinida, the Rhizopoda, and the Radiolaria. Some zoologists even have been anxious to relegate the sponges to the vegetable kingdom; but the botanists, who understood their business, refused to have anything to do with the intruders. And the botanists were quite right; for the discoveries of late years have not left the slightest doubt that the sponges are animal organisms, and animal organisms, too, of a very considerable amount of complexity, if we may regard as complex a structure which results from the building up and massing together of a number of similar parts.
The great majority of the sponges form a skeleton, which is composed of fibres of a horny texture, strengthened by needles, or spicula, of siliceous, or of calcareous, matter; and this framework is so connected together as to form a kind of fibrous skeleton. This, however, is not the essential part of the animal, [14] which is to be sought in that gelatinous substance, which invests the fibres of the skeleton during life, and is traversed by canals which open upon the surface of the sponge, directly or indirectly, by many minute, and fewer large, apertures.
If I may reduce a sponge to its simplest expression–taking the common Spongilla, for example, of our fresh waters,–the structure–removing all complexities, and not troubling ourselves with the skeleton, because that has nothing to do with what we are now considering–may be represented by the diagram (A, Fig. 4). There is a thin superficial layer (a) formed entirely of a number of the so-called sponge particles, or ultimate components of the living substance of the sponge, each of which is similar to an Amoeba, and contains a nucleus. These are all conjoined in a single layer, so as to form a continuous lamellar membrane, which constitutes the outer and superficial layer of the body. Beneath this is a wide cavity, communicating with the exterior by means of minute holes in the superficial layer (b), and, of course, filled with water. The cavity separates the superficial layer of the sponge from its deeper substance, which is of the same character as the superficial layer being made up of a number of aggregated sponge particles, each of which has a nucleus, and is competent to throw out numerous pseudopodial prolongations if detached. While the living sponge is contained in water, a great number of currents of water set in to the wide cavity beneath a, a, through the minute apertures (b), which have thence been termed "inhalent."
In the floor of the cavity there are a number of apertures which lead into canals ramifying in the deep layer, and eventually ending in the floors of certain comparatively lofty funnels, or craters. The top of each of these presents one of those larger and less numerous apertures, which have been referred to as existing on the surface of the sponge, and which are fitly termed "exhalent" apertures. For it has been discovered that strong, though minute, currents of water are constantly flowing out of these large apertures; being fed by the currents which as constantly set in, by the small apertures and through the superficial cavity, into the canals of the deeper substance. The cause of this very singular system of currents is the existence of vibratile
[15]
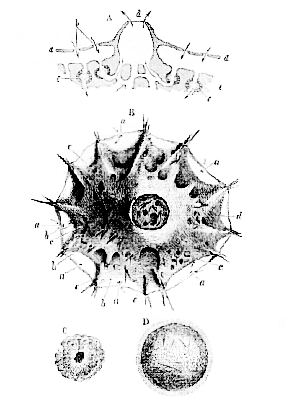
A, Hypothetical section of a Spongilla; a, superficial layer; b, inhalent aperture; c, ciliated chambers; d, an exhalent aperture; e, deeper substance of the sponge. The arrows indicate the direction of the currents. B, A small sponge with a single exhalent aperture, seen from above (after Lieberkühn); a, inhalent apertures; c, ciliated chambers; d, exhalent aperture. C, A ciliated chamber. D, A free-swimming ciliated embryo.
[16] cilia in the water-passages of the sponge, but it is only quite recently that the precise nature of the arrangement of the apparatus which gives rise to these currents, has been made out. The canals which enter the deep substance of the sponge become dilated into spheroidal chambers, lined with sponge particles (Fig. 4, A, c, C), each of which is provided with a vibratile cilium; and as all these cilia work in one direction–towards the crater–they sweep the water out in that direction, and its place is taken by fresh water, which flows in through the small apertures and through the superficial chamber. The currents of water carry along such matters as are suspended in them, and these are appropriated by the sponge particles lining the passages, in just the same way as any one of the Rhizopoda appropriates the particles of food it finds in the water about itself. So that we must not compare this system of apertures and canals to so many mouths and intestines; but the sponge represents a kind of subaqueous city, where the people are arranged about the streets and roads, in such a manner, that each can easily appropriate his food from the water as it passes along.
Two reproductive processes are known to occur in the sponges: the one of them, asexual, corresponding with the encysting process of the Gregarinida; and the other, truly sexual, and answering to the congress of the male and female elements in the higher animals. In the common fresh-water Spongilla, towards the autumn, the deeper layer of the sponge becomes full of exceedingly small bodies, sometimes called "seeds" or "gemmules," which are spheroidal, and have, at one point, an opening. Every one of these bags–in the walls of which are arranged a great number of very singular spicula, each resembling two toothed wheels joined by an axle–is, in point of fact, a mass of sponge particles which has set itself apart–gone into winter quarters, so to speak–and becoming quite quiescent, encysts itself and remains still. The whole Spongilla dies down, and the seeds, inclosed in their case, remain uninjured through the winter. When the spring arrives, the encysted masses within the "seed," stimulated by the altered temperature of the water, creep out of their nests, and straight[17]way grow up into Spongilla, like that from which they proceeded.
But there is, in addition, a true sexual process, which goes on during the summer months. Individual sponge particles become quiescent, and take on the character of ova; while, in other parts, particular sponge particles fill with granules, the latter eventually becoming converted into spermatozoa.
These sacs burst and some of the spermatozoa, coming into contact with the ova, impregnate them. The ova develop and grow into ciliated germs (D, Fig. 4), which make their way out, and, after swimming about for a while, settle themselves down and grow up into Spongillæ.
Now that we know the whole cycle of the life of the sponges, and the characters which may be demonstrated to be common to the whole of this important and remarkable class, I do not think any one who is acquainted with the organization or the functions of plants will be inclined to admit that the Spongida have the slightest real affinity with any division of the vegetable kingdom.
[27]
Notwithstanding that these animals differ very much in external appearance from the Polyzoa, we shall find a singular fundamental resemblance of internal structure between the two classes. All known Polyzoa are compound animals, that is to say, the product of every ovum gives rise, by gemmation, to great assemblages of partially independent organisms, or zooids. The Brachiopoda, on the contrary, are all simple, the product of each ovum not giving rise to others by gemmation. All the Brachiopoda possess a bivalve shell–a shell composed of two, more or less horny, or calcified, pieces, which are capable of a certain range of motion on one another, and are very commonly articulated together by teeth and sockets. The proper body, which is small when compared with the size of the shell, has its dorsal integument produced into broad membranous expansions, which line the interior of the valves of the shell, and are called the lobes of the mantle or "gallium." The aperture of the mouth is situated in the middle line, between the pallial lobes, and, on each side of it, is a longer or shorter prolongation of the body, provided with ciliated tentacula. It is from the presence of these "arms" that the class has received its name. The tentaculate oral disk of a Plumatella is already horse-shoe shaped (Figs. 8 and 9); suppose each crus of the horse-shoe to be pulled out to a much greater length, and [28] tentaculated "arms" would be produced, closely resembling those of the Brachiopoda.
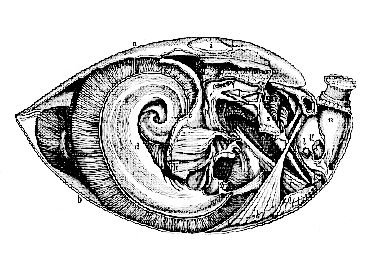
a, anterior layer of mantle; b, posterior layer; c. anterior walls of the body between the mantle lobes; d, arms; p, gullet; q, stomach, with cut biliary ducts of the left side; r, right hepatic mass; s, intestine ending cæcally between j and h; v, so-called "auricle" of the right "pseudo-heart," the left being almost wholly removed; w, pyriform vesicle fixed at the back of the stomach, and probably performing the function of a true heart; z, œsophageal ganglia.
The mouth leads into a gullet which is directed towards, or lies along, that side of the body from which one lobe of the mantle, the anterior, is continued; the gullet opens into a stomach, provided with a well-developed liver; and from the stomach, an intestine proceeds, which is directed towards, or [29] along, that side of the body from which the other lobe of the mantle proceeds; and then either ends, blindly, in the middle line (Fig. 10), or else terminates in a distinct anus between the pallial lobes.
The principal ganglionic mass is situated behind and below the mouth, in the recentering angle between the gullet and the rectum; in other words, the intestine has, as in the Polyzoa, a neural flexure (Fig. 10). In all Brachiopoda which have been carefully dissected a singular system of cavities and canals situated in the interior of the body, but in free communication with the surrounding medium, has been discovered. This, which I shall term the "atrial" system (from its close correspondence with the system of cavities, which has received the same name in the Ascidians), has been wrongly regarded as a part of the true vascular system, and the organs by which it is placed in communication with the exterior have been described as "hearts." There are sometimes two and sometimes four of these "pseudo-hearts," situated in that part of the body wall which helps to bound the pallial chamber. Each pseudo-heart is divided into a narrow, elongated, external portion (the so-called "ventricle"), which communicates, as Mr. Hancock has proved, by a small apical aperture with the pallial cavity; and a broad, funnel-shaped inner division (the so-called "auricle"), communicating on the one hand by a constricted neck with the so-called "ventricle," and, on the other, by a wide, patent mouth, with a chamber which occupies most of the cavity of the body proper, and sends more or less branched diverticula into the pallial lobes. These have been described as parts of the blood vascular system; and arterial trunks, which have no existence, have been imagined to connect the apices of the ventricles with vascular networks of a similarly mythical character, supposed to open into the branched diverticula.
In fact, as Mr. Hancock has so well shown in his splendid and exhaustive memoir, published in the Philosophical Transactions for 1857, the true vascular system is completely distinct from this remarkable series of "atrial" chambers and canals, the function of which would appear to be to convey away excretory matters and the products of the reproductive organs, [30] which are developed in various parts of the walls of the atrial system.
The precise characters of the true vascular system of the Brachiopoda probably require still further elaboration than they have yet received; and the same may be said, notwithstanding the valuable contributions of F. Müller and of Lacaze Duthiers, of their development; but the shell, the pallial lobes, the intestine, and the nervous and the atrial systems afford characters amply sufficient to define the class.
[42]
The star-fishes, sea-urchins, sea-cucumbers, trepangs, and feather-stars–known technically as Asteridea, Echinidea, Holothuridea, Ophiuridea, Crinoidea, &c., are marine animals which differ vastly in external appearance, though they all, in the adult state, present a more or less definitely radiate arrangement of some parts of their organization.
That which most remarkably distinguishes the Echinodermata is the nature of the embryo, and the strange character of the process by which the adult form is originated by a secondary development within that embryo.
In the great majority1 of the Echinodermata, the development of which has been examined, the impregnated egg gives rise to a free-swimming, ovoid, ciliated embryo, the cilia of [43] which soon become restricted to, and, at the same time, largely developed upon, one, two, or more bands, which are disposed either transversely, or more or less obliquely to the longitudinal axis of the body, but which are, in any case, bilaterally symmetrical (Fig. 18).
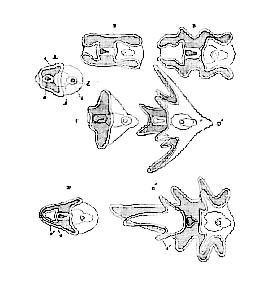
A. Common form whence the Holothurid (B, B') and Ophiurid or Echinid (C, C') larvæ are derived. D, D'. Younger and more advanced stages of the Asterid (Bipinnaria) larvæ. a. Mouth. b. Stomach. c. Intestine. d. Anus. e, Cilated band. e'. Second or anterior ciliated circlet.
The parts of the body which carry the ciliated band, or bands, often become developed into processes, which correspond upon each side of the body, and thus render its bilateral symmetry more marked (Fig. 18, C', D'). And, in the larvæ of some Echinidea and Ophiuridea, other bilaterally symmetrical processes are developed from parts of the body which do not lie in the course of the ciliated bands.
The larvæ of Asteridea and Holothuridea are devoid of any [44] continuous skeleton, but those of Ophiuridea and Echinidea possess a very remarkable bilaterally symmetrical, continuous, calcareous skeleton, which extends into, and supports the processes of the body (Fig. 21).
A distinctly defined alimentary canal early makes its appearance in these Echinoderm larvæ. It is divided into a well-marked oral and œsophageal portion, a globular stomach, and a short intestine terminating in an anal aperture (Figs. 18 and 19). All the parts of the alimentary canal are disposed in a longitudinal and vertical plane, dividing the larval body into two symmetrical halves; but the œsophageal and intestinal portions are so disposed as to make an angle, open towards the ventral side, with one another. No nervous, or other organs, besides those indicated, have as yet been discovered in these larvæ.
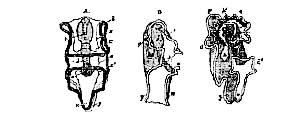
A. Ventral. B. Lateral view of the larva. C. Commencing rudiment of the starfish. a. Mouth. b. Œsophagus. c. Stomach. c'. Intestine. o. Anus. x. Anterior, and y, principal ciliated band. h. Cæcal diverticulum, forming the rudiment of the ambulacral vascular system, and opening externally by the pore, g. h'. Perisoma of the starfish.
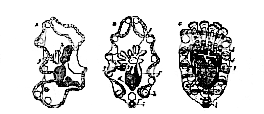
A. Early condition of larva. B. C. Later stages. f, g, h, the ambulacral vascular system.
[45]

A, earliest, and B. later condition of larva. C. The Echinid imago developed within and nearly obliterating the larva.
After swimming about in this condition for a while, the larva begins to show the first signs of those changes by which it is converted into the adult Echinoderm. An involution of the integument takes place upon one side of the dorsal region of the body, so as to give rise to a cæcal tube, which gradually elongates inwards, and eventually reaches a mass of formative matter, or blastema, aggregated upon one side of the stomach. Within this, the end of the tube becomes converted into a circular vessel, from which trunks pass off, radially, through the enlarging blastema. The latter, gradually expanding, gives rise in the Echinidea, the Asteridea, the Ophiuridea, and the Crinoidea, to the body-wall of the adult; the larval body and skeleton (when the latter exists), with more or less of the primitive intestine, being either cast off as a whole, or disappearing, or becoming incorporated with the secondary development, while a new mouth is developed in the centre of the ring formed by the circular vessel. The vessels which radiate from the latter give off diverticula to communicate with the cavities of numerous processes of the body–the so-called feet–which are the chief locomotive organs of the adult. The radiating and circular vessels, with all their appendages, constitute what is known as the "ambulacral system;" and, in Asterids and Echinids, this remarkable system of vessels remains in communication with the exterior of the body by canals, connected with perforated portions of the external skeleton–the so-called "madreporic canals" and "tubercles." In Ophiurids the persistence of any such communication of the ambulacral system [46] with the exterior is doubtful, and still more so in Crinoids. In Holothurids no such communication obtains, the madreporic canals and their tubercles depending freely from the circular canal into the perivisceral cavity.
Whether the larva possessed a skeleton or not, the adult Echinoderm presents a calcareous framework which is developed quite independently of that of the larva. This skeleton may be composed of mere detached spicula, or plates, as in the Holothurids; or of definitely disposed ossicula, or regular plates, as in other Echinoderms. In the latter case its parts are always disposed with a certain reference to the disposition of the ambulacral system, and hence have a more or less distinctly radiate arrangement. It might be expected, in fact, that the arrangement of the organs of support should follow more or less closely that of the chief organs of movement of the adult Echinoderm, and it is not surprising to find the nervous system similarly related. It is, in all adult Echinoderms, a ring-like, or polygonal, ganglionated cord, situated superficially to that part of the ambulacral system which surrounds the mouths and sending prolongations parallel with, and superficial to, the radiating ambulacral trunks.
The reproductive organs of the Echinoderms, which usually open upon, or between, parts of the radially disposed skeleton, commonly partake of the radial symmetry of that skeleton; but they have no such radial symmetry in the Holothuridea.
The alimentary canal of the adult Echinoderm is still less dependent upon the skeleton, and only in one group, the Asteridea, exhibits anything approaching a radiate disposition. Where skeletal elements are developed around the mouth or gullet, however, they have a radial disposition; as, e. g., the parts of the so-called "lantern of Aristotle."
The vascular system which exists in many, if not all, adult Echinoderms, but the true nature of which is by no means understood at present, is closely related both to the alimentary and to the ambulacral systems, and partakes of the disposition of both.
No Echinoderm whatsoever has its organs, internal or external, disposed with that absolute and perfect radial symmetry [47] which is exhibited by a Medusa, the tendency towards that kind of symmetry being always disturbed, either by the disposition of the alimentary canal, or by that of some part of the ambulacral apparatus. Very often, as in the Spatangoid sea-urchins, and in many Holothuridea, the ambulacral and nervous systems alone exhibit traces of a radial arrangement; and in the larval state, as we have seen, radial symmetry is totally absent, the young Echinoderm exhibiting as complete a bilateral symmetry as Annelids, or Insects.
1 In Ophilepis squamata and Echinaster sepositus, the larva appears to attain only a very imperfect state of development before the appearance of the echinoderm body; and careful re-examination is required to decide how far the larvæ of these animals are truly bilaterally symmetrical.
[55]
It is this extreme elasticity, if I may so speak, of the crustacean type which renders the construction of any definition of the Crustacea, which shall include all its members and exclude the present class, so difficult. For the Spiders, Scorpions, Mites, and Ticks, which constitute this class, possess all the characters which have been just stated to be common to the Crustacea save one; when they are provided with distinct respiratory organs, in fact, these are not external branchiæ, adapted for breathing ærated water, but are a sort of involution of the integument in the form of tracheal tubes, or pulmonary sacs, fitted for the breathing of air directly. But then many of the lower Arachnida, like the lower Crustacea, are devoid of special respiratory organs, and so the diagnostic character fails to be of service.
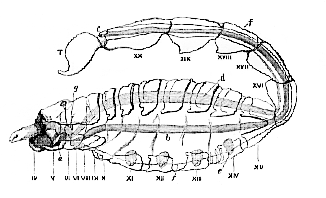
a, Mouth leading into the pharyngeal pump. The large labrum lies above the mouth, and at the side of it are the bases of the large chelæ, or mandibles, IV., and above them the cheliceræ, or antennæ. vi. to xx. Somites of the body. T. Nelson; b, intestine; c, anus; d, indicates the position of the heart. e, the pulmonary sacs; f, a line indicating the position of the ganglionic chain; g, the cerebral ganglia.
The following common characters of the Arachnida, how[56]ever, help out our diagnosis in practice. They never possess more than four pairs of locomotive limbs, and the somites of the abdomen, even when the latter is well developed, are not provided with limbs. Again, in the higher Arachnida (Fig. 28), as in the higher Crustacea, the body is composed of twenty somites, six of which are allotted to the head; but, in the former class, one of the two normal pairs of antennæ is never developed, and the eyes are always sessile, while, in the highest Crustacea, the eyes are mounted upon moveable peduncles, and both pairs of antennæ are developed.
|
THE
HUXLEY
FILE

|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
{kind=link}