[241] REVIEW I.
1. De Partibus Similaribus. By G. Fallopius. (Date uncertain, before 1562.)
2. Theoria Generationis. By Caspar Friederich Wolff. 1759.
3. Theorie von der Generation. By C. F. Wolff. 1764.
4. Theoria Generationis. By C. F. Wolff. Ed. Nova. 1774.
5. Von der eigenthümlichen und wesentlichen Kraft der Vegetablischen sowhol als auch der animalischen Substanz. By C. F. Wolff. 1789.
6. Enwickelungs Geschichte d. Thiere. By K. E. von Baer. 1828.
7. Untersuchungen über Phytogenesis. By Schleiden. 1837.
8. Mikroskopische Untersuchungun. By Schwann. 1838-9.
9. Ueber das Bindegewebe. By Reichert. 1845.
10. Die Vegetabilische Zelle. By H. von Mohl. 1850.
11. On the Mutual Relation of the Vital and Physical Forces. By W. B. Carpenter, M.D., F.R.S., &c, (‘Phil. Trans.’) 1850.
12 Handbuch der Gewebelehre. By A. Kolliker. 1852.
If we separate the elementary and essential facts of Life from all the various and complicated phenomena with which they are associated among the higher forms of living beings–if we examine those lowest and most rudimentary states of animal and vegetable existence, which are presented to us by the so-called ‘unicellular’ organisms, we find that the sole definable difference between a living thing and a mere formed morsel of some protein compound, fresh from the laboratory of the chemist is, that while the protein compound undergoes no change which may not be traced to the immediate and direct operation of some new or varying external condition, the Navicula or the Gregarina passes through the most remarkable successions of form, of size, and of chemical composition, which are equally definite in their nature, and equaloly certain in order of their occurrence, [242] whatever, within certain limits, be the change in external conditions, or whatever pains may be taken to prevent any variation in them.
Broadly, we may thus state the difference between the subjects of Physical and those of Biological Science: the former, the stone, the gas, and the crystal, have an inertia; they tend to remain as they are, unless some external influence affect them. The latter, animals and plants, on the other hand, are essentially characterized by the very opposite tendencies. As Reichert well expresses it:
"All organic bodies, therefore, represent, in relation to one another different and manifold states succeeding one another definitely upon a similar and homogeneous foundation; they form a common differential series, in which, independently of external conditions, a continual increase in the mutual differences and a diminution of the resemblances, occur." (p. 12.)
Linnæus seems to have wished to express his insight into this difference between living and dead matter, in his celebrated aphorism: "Stones grow; plants grow and live;" but so long as this "and live" was not analysed into its true meaning, the phrase marked the difference, but failed to define it.
Bichât recognised the independence of action of living beings in another way. All things which surround living beings tend, he says, to destroy them; but they nevertheless follow out their own appointed course. "La vie est l'ensemble des fonctions qui resistent à la mort."1
Now, this faculty of pursuing their own course, this inherent law of change, introduces, it will be observed, an element into the study of living beings which has no analogue in the world of ordinary matter. The latter frequently possesses structure, and may therefore be the legitimate subject of anatomy; but it undergoes no definite cyclical alterations,2 and, therefore, it offers nothing which corresponds [243] with what in living beings is called the History of Development, that branch of the investigation of structure which does not concern itself with the mere study of one state of a being–like anatomy–but examines into the manner in which the successive anatomical states are related to, and proceed out of, one another.
A profound physiologist and thinker, a contemporary and worthy rival of Haller, has beautifully expressed the relation of anatomy, of physiology, and of development (which he calls Generation), in the following words:
"The relations between anatomy, the doctrine of generation, and physiology, are about these. By anatomy we learn from observation the composition and structure of an organized body. We, however, are unable to explain this composition and structure; we only know that they are thus, and further than this we know nothing. But now, on the one hand, comes the doctrine of generation, in which that which we know from anatomy historically, is traced to its causes; on the other hand, we have physiology, in which the actions which an organized body is capable of producing are explained. Physiology is related to anatomy exactly as the corollary to the theorem from which it is deduced; my theory (of generation) is related to anatomy as its demonstration to the theorem."3
And again we find the relation of development to anatomy admirably and epigrammatically expressed in the 'Theoria Generationis' of the same writer. Development is, he says, "anatomia rationalis."
It has been said, and without doubt with profound truth, that the study of the structure of living beings originated in the wonder excited by their actions. But though this may, nay, must, have been the case at first, and though the curiosity of man has for three centuries past directed itself, with almost equal impartiality, to physiological, [244] anatomical, and developmental inquiries, still it is clear, that if the above account of the correlation of these branches of science be correct, their logical connexion, and the order, therefore, in which they must eventually arrive at perfection, is precisely the reverse. The striking and mysterious character of many of the functions may have led to the study of structure; but assuredly the understanding of the former presupposes a thorough knowledge of the latter.
It is conceivable that structure might be thoroughly made out without the least acquaintance with function, just as the ancient anatomists were well acquainted with the construction of the muscular system, and yet had no suspicion of its being the motor apparatus, and as at the present day we know full well the structure of the "vascular glands," though we can but guess their purpose; but it is quite impossible to attain to a complete knowledge of function without a thorough anatomical analysis. The action of the whole of any organ depends upon and is, that of the sum of its parts; it is, mechanically speaking, their resultant; so that until the nature and the precise modes of operation of all these parts have been made out, we can have no security that any law propounded concerning the functions of the whole, is other than a mere empirical generalization, liable to be interfered with at any moment, by the properties of some of the elementary parts with which we are unacquainted. Thus, up to within a few years ago, contractility was affirmed to be a general property of the cellular tissue of the skin; and this would have remained as an ascertained law, had not Kölliker shown, by the discovery of the extensive distribution of muscular fibre in it (that is, by pushing anatomical analysis a step further than it had previously been carried), that the supposed law was but an empirical generalization, and that the property of contractility, supposed to be inherent in the ordinary connective tissue of the skin, was, in fact, deducible from the presence of a totally different structure.
So again, Haller and his followers quoted the contraction of the heart, when removed from the body, as evidence of the innate contractility of muscle, apart from all nervous influence. This vis insita may exist or it may not, but further anatomical investigation has at any rate destroyed the force of the argument, by demonstrating the existence of nervous ganglia within the substance of the organ.
But enough of illustration of what must be sufficiently plain to any one who will reflect upon the subject; namely, that however much might be done towards the establishment of broad physiological truths, while the knowledge of structure was in a rough and imperfect state, still an exhaustive study of structure is absolutely necessary, [245] before any successful attempt can be made to establish the true laws of function, or to build the science of physiology upon an exact foundation.
Herein lies, consciously or unconsciously to their authors–for the man of genius is such, in virtue of having true and just tendencies and impulses, of which he often can give himself no logical account–the secret of the repeated attempts which have been made from the time of the very fathers of biology, to found what we now call the doctrine of general anatomy or histology, which is, in other words, the exhaustive anatomical analysis of organized bodies. That animals and plants, complex as they may appear, are yet composed of comparatively few elementary parts, frequently repeated, had been noticed by the profound intellect of Aristotle; and Fallopius tells us that Galen had attained to still more clear and definite conceptions with regard to these "partes similares" or "simplices".4
"Galenus per simplices partes eas intelligit quæ non constant ex dissimilibus substantiis, in quas resolvitur corpus humanum, nec ultra datur progressio et istæ partes dicuntur simplices quia cum ad hoc ventum fuerit in resolutione corporis humani, non amplius progredi possumus." (p. 103.)
Such, indeed, must be the definition of elementary parts at all periods of science–they are ultimate, because we can go no further; though it is of course a very different matter whether we are stopped by the imperfection of our instruments of analysis, as these older observers were, or by having really arrived at parts no longer analyzable.
The celebrated professor of Modena, whose words we have just cited, was one of the first of those who carried the light shed by the revival of letters into the region of medicine and its allied sciences; and his work 'De Partibus Similaribus,' from which they are taken, must excite the admiration of every modern reader, not merely by the critical acumen and original genius which it displays, but by the scientific and absolutely accurate manner in which the whole subject of general anatomy is handled.
The classes of "partes similares," or tissues, of which he treats, are bone, cartilage, fat, flesh, nerve, ligament, tendon, membrane, vein, artery, nails, hairs, and skin; and he examines and details under each head the minute structure, so far as it was accessible to his means of investigation; the chemical and physical properties (expressed, of course, in the language of the day), and even the peculiarities [246] manifested by the diseased state. Nor is he at all wanting in what has been considered, and justly, to be Bichât's great merit–an essentially positive method of studying the tissues, inasmuch as he particularly insists on the necessity of investigating the properties of each tissue for itself, and of avoiding all hypothetical speculation; in fact, with the quaint plainness of the age, he does not hesitate to insinuate that Averrhöes must have been "ebrius" when he discoursed touching "spiritus qui insensibiles sunt."
The vitality of each tissue, independently of every influence save the general conditions of nutrition, is maintained by Fallopius, not as a mere speculation, but on sound embryological grounds. How can the liver, he asks, be the sole source and prime mover of all vital organization, as some have maintained, when, in the development of the chick, we see other organs appear before it? All that the liver and the vessels can do is to modify the supplies, by affecting the "restitutiones spirituum ac nutrimentis" (p. 98), the "partes similares" themselves having a "regimen insitum," or, as in our day it would be called, "vital force," of their own; and he quotes, as expressing his own views, the following remarkable passage from Actuarius:
"Quod partes naturales agunt propriâ formâ ac cum instrumento quod dicitur spiritus animalis: nam hoc instrumento per propriam formam attrahunt, concoquunt et expellunt, et hic spiritus est immediatum instrumentum vis naturalis, et hic spiritus dicit Actuarius, originem ducit unâ cum formâ ipsius particulæ, et ex eâdem materiâ eodemque tempore fit."
Substitute here for the indefinite "particulæ" definite vesicular particles or cells, and for "spiritus animalis" the modern terms of equivalent meaning or no meaning–vital-force or cell-force, and this passage would serve very well as a concise expression of the "cell-theory," such as may be found in many a hand-book of the day. So far, and no further, have three centuries brought us!
In fact, it must be confessed, that these old writers were fully possessed (more so, in truth, than many of their successors) with the two fundamental notions of structural and physiological biology; the first, that living beings may be resolved anatomically into a comparatively small number of simple structural elements; the second, that these elementary parts possess vital properties, which depend for their manifestation only upon the existence of certain general conditions (supply of proper nutriment, &c.), and are independent of all direct influence from other parts.
But it would seem, that Truth must pass through more than one Avatar, before she can attain a firm hold upon the mind even of men [247] of science–and at the end of the eighteenth century it required all the genius of Bichât to sift the wheat from the chaff, amongst the great mass of facts which the observation of the past ages had accumulated–and strengthening whatever place was weakest by new investigations–to establish these very two propositions, upon a broad and henceforward firm foundation. Great as was the service which Bichât rendered in this way to biology–and wide as the difference between the treatise 'De Partibus Similaribus' and the 'Anatomie Generale' may be–still the one is the intellectual progeny of the other, and exhibits neither alteration nor improvement in the method pursued.
In the meanwhile, however, an aid to investigation had arisen, by the means of which this method could be pushed to its uttermost limits–we refer to the invention of the microscope. The influence of this mighty instrument of research upon biology, can only be compared to that of the galvanic battery, in the hands of Davy, upon chemistry. It has enabled proximate analysis to become ultimate. Without the microscope the ultimate histological elements were, as we have seen, defined negatively, as parts in which any further structural difference was too small to be detected. The microscope, on the other hand, enables us to define the tissues positively–to say, a given tissue has such a structure, and magnify it as you will, it will present no further differences.
The amount of such positive information as to the ultimate structure of the tissues, collected by Leuwenhoek, Malpighi, and their successors, between the middle of the seventeenth and the fourth decade of the present century, was very great, and in fact the most important and characteristic features presented by the histological element of plants and animals may be said to have been well made out, at the time of the appearance of the celebrated treatises by Schleiden and Schwann, cited at the head of this article; and these writers, therefore, added but little to the body of knowledge in this direction. It is most unquestionable, however, that the biological sciences, and more especially histology, received a wonderful stimulus at their hands. Whatever cavillers may say, it is certain that histology before 1838 and histology since then, are two different sciences–in scope, in purpose, and in dignity–and the eminent men to whom we allude, may safely answer all detraction by a proud "circumspice."
But wherein does the real value of their work lie? We think this question may be readily enough answered by those who admit the force of what has been said in our opening paragraphs–who acknowledge that mere anatomy does not exhaust the structure of living beings–and that before histology can be said to be complete, we must [248] have a histological development as well as a histological anatomy. Leuwenhoek and the majority of his successors had enough to do in making out the "historicam cognitionem," the simple anatomy of the tissues; it tasked all their powers to arrive at a clear statement of the "theorem," while it is the great merit of Schleiden and Schwann that they sought to arrive at an "anatomia rationalis," and to furnish the "demonstration of the theorem." The old method of investigation had been carried as far as it would go, and they applied the only other which remained, and made it familiar to the general mind. Turn to any of Schleiden's works, and we find the logical acuteness, and the vituperative sarcasm, which he wields with equal force, employed in urging the study of development as the one thing needful for scientific botany. And Schwann's entire essay testifies to what he expressly tells the reader, that his investigations are distinguished from all others by being based upon the study of development. Let one citation suffice:–"The theory of the present investigation was, therefore, to show .... that there exists a common principle of development for all the elementary parts of the organism." (Schwann, pp. 193–196.)
Intending as we do to venture upon a critical examination of the absolute value of Schleiden's and Schwann's contributions to biological science, which may lead us to conclusions not ordinarily admitted, we have been particularly desirous to estimate fairly the position which they occupy in its history, and the influence which their labours have had upon its progress–which is a widely different matter–for, in attempting to weigh the labours of others, we should be in danger of committing great injustice, if we did not carefully bear in mind that paradoxical as it may seem, the value of a theory and its truth, are by no means commensurate. In so complex a science as that which relates to living beings, accurate and diligent empirical observation, though the best of things as far as it goes, will not take us very far; and the mere accumulation of facts without generalization and classification is as great an error intellectually, as, hygienically, would be the attempt to strengthen by accumulating nourishment without due attention to the primæ viæ, the result in each case being chiefly giddiness and confusion in the head.
In biology, as in all the more complicated branches of inquiry, progress can only be made by a careful combination of the deductive method with the inductive, and by bringing the powerful aid of the imagination, kept, of course, in due and rigid subordination, to assist the faculties of observation and reasoning; and there are periods in the history of every science when a false hypothesis is not only better [249] than none at all, but is a necessary forerunner of, and preparation for, the true one. As Schwann himself well expresses it:
"An hypothesis is never hurtful, so long as one bears in mind the amount of its probability, and the grounds upon which it is formed. It is not only advantageous, but necessary to science, that when a certain cycle of phenomena have been ascertained by observation, some provisional explanation should be devised as closely as possible in accordance with them; even though there be a risk of upsetting this explanation by further investigation; for it is only in this way that one can rationally be led to new discoveries, which may either confirm or refute it." (p. 221.)
The value of an hypothesis may, in fact, be said to be twofold–to the original investigator, its worth consists more in what it suggests than in what it teaches; let it be enunciated with perspicuity, so that its logical consequences may be clearly deduced, and made the base of definite questions to nature–questions to which she must answer yes or no–and of its absolute truth or falsehood, he recks little: for the mass of men, again, who can afford no time for original research, and for the worker himself, so far as respects subjects with which he is not immediately occupied, some system of artificial memory is absolutely necessary. This want is supplied by some "appropriate conception" which, as Dr. Whewell would say, "colligates" the facts–ties them up in bundles ready to hand–by some hypothesis, in short. Doubtless the truer a theory is,–the more "appropriate" the colligating conception,–the better will it serve its mnemonic purpose, but its absolute truth is neither necessary to its usefulness, nor indeed in any way cognizable by the human faculties. Now it appears to us that Schwann and Schleiden have performed precisely this service to the biological sciences. At a time when the researches of innumerable guideless investigators, called into existence by the tempting facilities offered by the improvement of microscopes, threatened to swamp science in minutiæ, and to render the noble calling of the physiologist identical with that of the 'putter-up' of preparations, they stepped forward with the cell-theory as a colligation of the facts. To the investigator, they afforded a clear basis and starting-point for his inquiries; for the student, they grouped together immense masses of details in a clear and perspicuous manner. Let us not be ungrateful for what they brought. If not absolutely true, it was the truest thing that had been done in biology for half a century.
But who seeks for absolute truth? Flattering as they were to our vanity, we fear it must be confessed that the days of the high a priori road are over. Men of science have given up the attempt to soar [250] eagle-like to some point amidst the clouds, whence the absolute relations of things could be securely viewed; and at present, their more useful, if more ignoble course, may rather be compared to that of the flocks of sparrows in autumn, which one sees continually halting yet always advancing–flying from tree to tree, noisily jubilating in each, as if that were assuredly the final resting-place and secure haven of sparrows, and yet as certainly taking their departure after awhile, in search of new acquisitions. We must build our theories, in these days, as we do our houses: giving up all attempt at Cyclopean architecture, let us bethink ourselves rather of the convenience of our successors, who will assuredly alter, and perhaps pull them down, to suit the needs of their own age; and if we seek their gratitude, let us strive not so much to knit our materials firmly together (which will only give them more trouble and yield us less thanks), as to see that they are separately sound and convertible. This much digression has seemed necessary, by way of securing ourselves from any suspicion of a desire to under-estimate the historical value of Schleiden and Schwann's researches, in the course of an attempt to show that they are based upon errors in anatomy, and lead to errors in physiology.
Again, with regard to that value, we have a few words to say in a merely historical point of view. The sketch we have given of the progress of general anatomy, we believe, omits mention of no ordinarily recognized epoch, nor fails to indicate the acknowledged order of the successive introduction of those great leading ideas with which we are at present concerned.5 In their own belief, and in that of their contemporaries, Schleiden and Schwann have not only worked out developmental histology, but originated it; and the latter, in his reclamation against Valentin (loc. cit., pp. 260, 261), defends his claim to be considered the originator of the idea that "a common principle governs the development of the elementary parts of all organisms." Now, we fully recognise the originality of these writers; we believe that they deserve all the credit which can attach to a noble plan carried out with no small success; and we further remember that the majority always sympathizes with the cry, "Pereant qui ante nos, nostra," &c.; but, as we have said, truth often has more than one Avatar, and whatever the forgetfulness of men, history should be just, and not allow those who had the misfortune to be before their time, to pass for that reason into oblivion.
Such was the position into which his great genius forced Caspar [251] Friedrich Wolff–such the fate with which he has met. The manuals of physiology tell us that he was the founder of the doctrine of epigenesis–a doctrine which, in the present day, seems so plain and obvious, that we do not give him much credit for it, forgetting that he had to struggle against the authority of Malpighi and of Haller, and the attacks of Bonnet; influence and authority so great, that though every reader of the 'Theoria Generationis' must see that Wolff triumphantly establishes his position, yet, seventy years afterwards, we find even Cuvier6 still accrediting the doctrine of his opponents.
It is less generally (we might say hardly at all) known that Wolff demonstrated, by numerous observations on development, the doctrine of the metamorphosis of plants, when Göthe, to whom it is commonly ascribed, was not quite ten years old;7 but it seems to have been wholly forgotten that he endeavoured to work out, upon the basis of the strict study of histological and morphological development, that "identity of structure of plants and animals" which is the thesis defended by Schwann. Had Wolff's teaching been founded upon one of those clever guesses upon which an able man will often build up a plausible hypothesis, we should have thought it quite unnecessary to make even historical reference to him; but the most cursory examination of the 'Theoria Generationis,' or of the more popular and discursive exposition of his views in the 'Theorie von d. Generation,' is enough to dispel any such notion. The passage we have already quoted is sufficient to show how just and accurate Wolff's ideas upon the importance of the study of development, as a method, were; and the whole of his work is the laborious application of that method. The parts of the calyx, of the corolla, and of the pericarp, are for him "modified leaves;" not because certain observed modifications had suggested that they might be so considered–which is the whole gist of Göthe's subsequent argument–but because he had carefully traced back their development, and had found that they all proceeded from the same original form. The homology of the wing of the chick with its leg is placed by Wolff on precisely the same basis–the only one, be it observed, on which any homology can ultimately rest; and following out the argument to its legitimate conclusion, he shows that the appendicular organs of plants and animals are developed after the [252] same fashion. The limbs of animals, he says, are developed in the same manner from the body of the embryo, as the leaf from the stem, or the lamina of the leaf from its mid-rib. Ordinary four-footed animals are like pinnatifid leaves, while "the bat is a perfect leaf–a startling statement, but, as I have shown, the analogy is not chimerical for the mode of origin of the two is the same."8
Wolff's doctrine concerning histological development is shortly this.9 Every organ, he says, is composed at first of a little mass of clear, viscous, nutritive fluid, which possesses no organization of any kind, but is at most composed of globules. In this semi-fluid mass, cavities (Bläschen, Zellen) are now developed; these, if they remain rounded or polygonal, become the subsequent cells–if they elongate, the vessels; and the process is identically the same, whether it is examined in the vegetating point of a plant, or in the young budding organs of an animal. Both cells and vessels may subsequently be thickened, by deposits from the "solidescible" nutritive fluid. In the plant, the cells at first communicate, but subsequently become separated from one another; in the animal, they always remain in communication. In each case, they are mere cavities, and not independent entities; organization is not effected by them, but they are the visible results of the action of the organizing power inherent in the living mass, or what Wolff calls the vis essentialis. For him, however, this "vis essentialis" is no mythical archæus, but simply a convenient name for two facts which he takes a great deal of trouble to demonstrate; the first, the existence in living tissues (before any passages are developed in them) of currents of the nutritious fluid determined to particular parts by some power which is independent of all external influence; and the second, the peculiar changes of form and composition, which take place in the same manner.
Now there is really no very great difference between these views of the mode of development of the tissues and those of Schleiden and Schwann. The "solidescible nutritive fluid" of Wolff is the "cytoblastema" of Schleiden and Schwann; with the exception of the supposed relation of the nucleus to the development of the cell (which, as we shall see, is incorrect) Wolff's description of the latter process is nearly that of Schleiden; Wolff maintains that the "vessels" of plants are the result of the greater activity of the nutritive currents in particular directions; and so does Schleiden.10
[253] Examining his statements closely, we notice, indeed, that his imperfect means of investigation led Wolff into two important errors–that of supposing the cells of plants to communicate in their youngest state, and thence deducing a false analogy with the areolar tissue of animals; and that of supposing that animal and vegetable tissues are always, in their very youngest state, absolutely structureless. However, as we shall see subsequently, Wolff is by no means singular in having started with grave anatomical mistakes, and we cannot perceive that in his case these errors, one of which, at any rate, Schleiden shares with him, vitiate those other and more important parts of his views, to which we are about to refer.
We have said, in fact, that not merely speculatively, but by observation, Wolff established a theory of the development of the vegetable tissues very similar to that of Schleiden, and that "identity of structure as shown by their development," between plants and animals, to prove which, was the purpose of the microscopical investigations of Schwann. But he did much more than this. In the 'Theoria Generationis,' and in the essay on the vital forces published thirty years afterwards, Wolff developed some very remarkable views on the relation of life to organization–of the vital processes to the organic elements–in which he diverges very widely from all who preceded and from most who have followed him, most of all from Schleiden and Schwann. We may best exhibit the bearing of these views by contrasting them with those of the latter writers.
Schleiden and Schwann teach implicitly that the primary histological elements (cells) are independent, anatomically anal physiologically; that they stand in the relation of causes or centres to organization and the "organizing force;" and that the whole organism is the result of [254] the union and combined action of these primarily separate elements. Wolff, on the other hand, asserts that the primary histological elements (cells too, but not always defined in the same way) are not either anatomically or physiologically independent; that they stand in the relation of effects to the organizing or vital force (vis essentialis); and that the organism results from the "differentiation" of a primarily homogeneous whole into these parts. Such a doctrine is, in fact, a most obvious and almost a necessary development of the doctrine of epigenesis in general. To one who had worked out the conclusion, that the most complex, grosser, animal or vegetable organizations, arise from a semi-fluid and homogeneous mass, by the continual and successive establishment of differences in it, it would be only natural to suppose that the method of nature, in that finer organization which we call histological, was the same; and that as the organ is developed by the differentiation of cells, so the cells are the result of the differentiation of inorganic matter. If the organism be not constituted by the coalescence of its organs and tissues in consequence of their peculiar forces, but if, on the other hand, the organism exists before its organs and tissues, and evolves them from itself,–is it not probable that the organs and tissues also, are not produced by the coalescence of the cells of which they are composed, in consequence of their peculiar forces, but, contrariwise, that the cells are a product of the differentiation of something which existed before them?
For Schwann the organism is a beehive, its actions and forces resulting from the separate but harmonious action of all its parts (compare Schwann, l.c., p. 229). For Wolff it is a mosaic, every portion of which expresses only the conditions under which the formative power acted and the tendencies by which it was guided.
We have said above, not without a full consciousness of the responsibility of the assertion, that we believe the cell-theory of Schleiden and Schwann to be based upon erroneous conceptions of structure, and to lead to errors in physiology, and we beg now to offer some evidence in favour of these views. We need not stop to prove, what must be familiar to every one who is acquainted with Schwann's work, that in making his comparison of animal with vegetable structures, he rests wholly upon Schleiden's statements concerning the development, and upon the commonly prevalent views with respect to the anatomy, of the latter.
It is clear, then, that however logically consequent Schwann's work may be in itself, its truth and the justice of its nomenclature will depend upon that of these latter views and statements. Schwann took these [255] for granted, and if they were untrue he has been trusting to a rotten reed. Such, we fear, has indeed been the case. Schwann's botanical data were:
1. The prevalent notion of the anatomical independence of the vegetable cell, considered as a separate entity.
2. The prevalent conception of the structure of the vegetable cell.
3. The doctrine of the mode of its development.
Each of these, as assumed by Schwann, and as taught by Schleiden, has since, we shall endeavour to show, been proved to be erroneous. We will take them seriatim.
1. The first observer who, aided by the microscope, turned his attention to the structure of plants, was the versatile Hooke, and, as might be expected, the most noticeable thing to his mind was the existence of the innumerable cavities or "cells" scattered through their substance. Malpighi, the first proper botanical histologist, found that the walls of these vesicles were separable, that they could be isolated from one another, and therefore, doubtless urged more by the obvious convenience of the phraseology, than by any philosophical consideration upon the subject, he gave each the definite name of "utriculus," and regarded it as an independent entity. Of course it was a natural consequence that the plant should be regarded as constituted by the union and coalescence of a great number of these entities.
Grew, who if all scandal be true, is so much indebted to Malpighi, did not appropriate this view among other things; on the other hand, he compared the utricles to the cavities in the foam of beer; and subsequently Wolff propounded the idea, that the cells were cavities in a homogeneous substance, as we have mentioned above. In modern times, the most important defender of this mode of regarding the matter has been Mirbel, who (escaping the error of Wolff, that the cavities of the cells communicate) endeavoured to demonstrate its truth, by tracing the formation of the cambium; but, at the time when Schwann wrote, it must be considered to have been wholly discredited, the opposite view having one of its strongest supporters in the caustic Schleiden himself–as, indeed, would necessarily be the case, from the tendency of his researches upon phytogenesis. As we shall see below, however, Schleiden was quite wrong in his ideas of cell-development–and we have therefore merely to consider the purely anatomical arguments for the independence of the cell. Now these amount, however various their disguise, to nothing more than this–that, by certain chemical or mechanical means, a plant may be broken up into vesicles corresponding with the cavities which previously existed in it: of [256] course no one denies this fact; but of what value is it? Is the fact that a rhombohedron of calcareous spar breaks up, if pounded, into minute rhombohedrons, any evidence that those minuter ones were once independent, and formed the larger by their coalescence? Is the circumstance that wood itself tears up into fibres, any evidence that it was formed by the coalescence of fibres? Assuredly not; for every hand-book will tell us that these fibres are the result of a metamorphosis of quite different parts. Is it not perfectly clear, that the behaviour of a body under mechanical or chemical influences, is simply an evidence of the disposition of the lines of greatest cohesion or affinity among its particles at the time being, and bears not in the slightest degree upon the question as to what these lines indicate; whether they are the remains of an ancient separation among heterogeneous parts, or the expression of a recent separation which has arisen in a homogeneous whole? So that, if the walls of the cells were really, as distinct from one another as is commonly supposed, it would be no argument for their vital independence: but they are not so. Von Mohl has shown that, in the great majority of cases, the assumption of the existence of a so-called intercellular substance, depends simply on imperfect chemical investigation, that there exists no real line of demarcation between one cell and another, and that wherever cells have been separated, whether mechanically or chemically, there is evidence that the continuous cellulose substance has been torn or in some way destroyed. In young tissues–such, for instance, as the cambium, or the base of a leaf, we have been quite unable to detect the least evidence of the existence of any line of demarcation between the cells; the cellulose substance forms a partition between cavity and cavity, which becomes evenly blue throughout by the action of sulphuric acid and iodine, and which certainly, even under the highest powers, exhibits no symptom of any optical difference; so that, in this state, vegetable tissue answers pretty closely to Wolff's idea. It is a homogeneous cellulose-yielding, transparent substance, containing cavities, in which lie peculiar vesicular bodies, into whose composition much nitrogen enters. It will be found a great aid if in the present confused state of terminology the reader will accept two new denominations for these elementary parts, which express nothing but their mutual relation. To the former, and to everything which answers to it, we shall throughout the present article give the name of Periplast, or periplastic substance; to the latter, that of Endoplast. So far, then, from the utricles or cells in the plant being anatomically distinct, we regard it as quite certain that that portion which corresponds with the periplast, forms a continuous whole through the entire plant.
[257] 2. In 1837-8, each utricle of the plant was considered to have the following composition. In the first place, there was the cellulose cell-wall, or the portion of periplast answering to any particular endoplast; secondly, there were the cell-contents, a substance of not very defined nature, which occupied the cavity of the cell; and thirdly, there was the nucleus, a body to whose occurrence attention was first drawn, as is well known, by our own illustrious botanist, Robert Brown. He, however, cautiously remarked only its very general occurrence, without pretending to draw any inference from the fact; while Schleiden made the belief in its existence in all young tissues, the first article of the faith botanical. This is, however, most certainly incorrect; there is no trace of a nucleus in many Algæ, such as Hydrodictyon, Vaucheria,11 Caulerpa; in the leaf of Sphagnum, nor in young germinating Ferns.12
Whatever opinion may be entertained upon this head, there is one point quite certain–the enumeration of the elements of the vegetable-cell given above is incomplete; there being one, and that the most important, which is omitted. We refer to the primordial utricle, which was only discovered by Von Mohl in 1844. This is a nitrogenous membrane, which always lies in close contact with the periplast, and forms, in fact, an included vesicle, within which the "contents" and the nucleus lie. Instead, therefore of the endoplast consisting merely of contents and a nucleus, it is a vesicle containing the two latter, when they exist at all; and they are of subordinate importance, for while, as we have seen, a nucleus and formed contents may be absent in young or even fully formed tissues, the primordial utricle is invariably present in the young structures, and often persists until they have attained their full size. Since, then, the functions of the vegetable "cell" can be effectually carried on by the primordial utricle alone; since the "nucleus" has precisely the same chemical composition as the primordial utricle; and since, in some cases of cell-division, new nuclei are seen to arise in the substance of the endoplast, by a mere process of chemical and morphological differentiation (Von Mohl, l. c., p. 52), it follows, we think, that the primordial utricle must be regarded as the essential part of the endoplast–the protoplasm and nucleus being simply its subordinate, and, we had almost said, accidental anatomical modifications.
3. Finally, with respect to Schleiden's observations upon the mode of cell-development, according to which in all cases the new production of vegetable-cells takes place by the development of nuclei, round [258] which the cell-membrane is deposited, subsequently expanding and becoming separated from the nucleus, so as to form a complete cell; we need only say, that they have been long since set aside by the common consent of all observers; in Von Mohl's words (p. 59): "The whole of this account of the relation of the nucleus to the cell-membrane is incorrect." The fact is, that in by far the greater proportion of cases, new cell-development occurs by the division of the previous endoplasts, and the growth or deposition round them and between them, of fresh periplastic substance. The extent of this process of division will be understood, if we remember that all observers now agree in its being the method by which "celldevelopment" always occurs, except in the embryo-sac of the Phanerogamia, the sporangia of Lichens and of some Algæ and Fungi. The so-called free cell-development of the latter, however, by no means takes place in accordance with Schleiden's views, but by the development of a cellulose membrane (periplast) around a mass of nitrogenous substance (endoplast), which may or may not contain a nucleus; subsequently increasing, pari passu, with the periplast. And it is well worthy of consideration, how far the process deserves any distinction, except in degree, from ordinary cell-division, since the new endoplast is only one portion of that of the parent cell, set aside for the purpose of fresh development, while the rest undergoes no corresponding change. However this may be, it may be regarded as quite certain that, leaving out of view the immediate results of sexual reproduction, the whole of the "cells,"–the entirety of the periplasts and endoplasts–of which a plant, whether it be a moss or an oak, are composed, never are independent of one another, and never have been so, at any period of their existence; but that, while the original endoplast of the embryo-cell, from which the plant sprung, has grown and divided into all the endoplasts of the adult, the original periplast has grown at a corresponding rate, and has formed one continuous and connected envelope from the very first. The ground of his comparison, therefore, is cut away from under Schwann's feet; every statement of Schleiden's on which he relied turning out to be erroneous–as we shall see if we turn to his original comparison of cartilage with a vegetable tissue (pp. 9–17). Schwann, finding in cartilage cavities with more or less distinct walls, in each of which lay a corpuscle, singularly resembling the nucleus of the vegetable-cell; finding also that the cell-wall was close to this corpuscle in the younger parts, more distant from it in the older (p. 24), naturally concluded that he had here, in the animal world, an exact confirmation of Schleiden's supposed discoveries, and of course gave to the corpuscle of cartilage the name of "cytoblast," [259] or "nucleus," as indicating its homology with the structure of that name, in the plant.
The primordial utricle was, as we have said, not then discovered in the latter, and of course Schwann was not led to look for anything corresponding to it. Indeed, had he done so, his search would have been unsuccessful, for the young and unaltered cartilage cavity contains the corpuscle, and nothing else. The circumstance, therefore, which Schwann considered to demonstrate the identity of structure of plants and animals–i.e., the correspondence of the cartilage-corpuscle with the nucleus of the vegetable -cell, and of the chondrin-wall with the cellulose-wall, would, if it were really the case, be the widest possible ground of distinction between the two, for it would leave the most important element of the latter, the "primordial utricle," without any homologue in the animal, and totally unaccounted for.
Fig. 1. A, Collenchyma cells of Beta vulgaris; B, Stellate, tissue of the pith of the rush; C, a cartilage-cell with its corpuscle, to compare with D, a vegetable-cell with its nucleus, the primordial utricle in the latter being indicated only by a dotted line.
It is precisely the neglect of this important change in the whole subject, effected by the discovery of Von Mohl, which has, we think, led to the confusion which prevails at present, not only in the comparative, but in the absolute nomenclature of animal histology. Animal physiologists go on using Schwann's nomenclature, forgetting that the whole doctrine of the vegetable-cell, from which he drew that nomenclature, has been completely upset; and at present, beyond the mere fact of a common vesicularity at one period of their existence, one would be led, on opening successively two works on animal and vegetable structure, rather to predicate their total discrepancy, than any uniformity between them.
Now does this discrepancy lie in the facts, or in our names of them? To decide this question, it seemed to us that the only plan was to follow Schwann's steps, and to compare cartilage with a vegetable tissue–for he has shown logically and conclusively enough, that whatever is true of the corpuscle of cartilage, to which he gives the name of "nucleus," is true also of all those corpuscles in the other tissues, to which he gives the same name.
[260] Let us compare, then, some young vegetable tissue, say that of the base of a Sphagnum leaf (fig. 2, A), which is in many respects very convenient for examination, with that of young cartilage (fig. 3, A); the identity of structure is such, that it would be difficult, without the aid of chemical re-agents, to distinguish one from the other: in each, we see highly nitrogenous, more or less vesicular endoplasts imbedded in a homogeneous transparent substance, whose cavities they wholly fill. If we trace the further development, we find that in the Sphagnum leaf the endoplasts and their cavities rapidly increase in size (fig. 2, B), the former becoming, in certain localities of the leaf, regular primordial utricles without any nucleus, and growing in exact proportion to the cavities in the periplast (b), while in other directions, having attained a certain size they cease to grow, and rapidly disappear, leaving the periplastic cavity empty (a). In cartilage precisely the same thing occurs. The endoplasts increase in size for awhile, and then stop, while the periplastic cavities continue to increase, and thus we have eventually a cartilage-cavity with its corpuscle. In old cartilage the latter frequently disappears, or is converted into fat. We have here purposely selected, in both the animal and the plant, simple cases, in which the endoplast becomes a primordial utricle, without any nucleus. Had we selected the cambium of a phœnogamous plant, it would have been merely necessary to add that, as the endoplast grew, a nucleus appeared in its interior; and in ossifying cartilage, near the ossifying surface, we have repeatedly seen endoplasts such as those described above, some of which contained definite "nuclei," while those in their immediate neighbourhood possessed none.
In the case of cartilage, then (and it is a conclusion at which Leidy and Remak have already arrived), we hold it to be proved that the corpuscle does not correspond with the nucleus of the plant as Schwann supposed, but with the primordial utricle, contents and nucleus; or, in other words, that the "nucleus" of cartilage is the equivalent of the "primordial utricle" of the plant–that they are both endoplasts. It follows, hence, that the chondrin-wall of the cartilage is the homologue of the cellulose wall of the plant, and that they both represent the periplastic element. The phenomena of growth and multiplication exhibited by these corresponding elements are perfectly similar. The process of cell-division, as it is called, is identical in each case. In the plant, the primordial utricles divide, separate, and the cellulose substance grows in between the two. In young cartilage the same thing occurs, the corpuscles divide, separate, and the chondrin substance eventually forms a wall of separation between the two. There is neither endogenous development nor new formation in either case. [261] The endoplasts grow and divide, the periplast grows so as to surround the endoplasts completely, and except so far as its tendency is, to fill up the space left by their separation, there is no evidence that its growth is in any way affected by them, still less, that it is, as is often assumed, deposited by them. We are led, then, to the conclusion that though Schwann's great principle of the identity of structure of plants and animals is perfectly correct, his exposition of it is incorrect, inasmuch as the corpuscle of cartilage (his "nucleus," whence he reasoned to the other "nuclei") answers not to the "nucleus," but to the "primordial utricle" of the plant; since the mode of development of new "cells," though identical in each case, is different from what Schleiden stated, and Schwann believed; and, finally, since, for the notion of the anatomical independence of the cells, we must substitute that of the unity and continuity of the periplastic substance in each case.
Intimately connected with these structural errors, as we cannot but think them, are Schwann's views of the nature and powers of the "cell," and those subsequently developed (principally by Kölliker) with respect to the action of the nuclei as "centres of force." Led apparently by his views of its anatomical independence, Schwann maintains, as a general proposition, that the cell as such possesses powers which are not inherent in its separate molecules.
"We must, in fact, ascribe an independent life to cells–i.e., the combination of molecules which take place in a single cell are sufficient to set free the force, in consequence of which the cell has the power of attracting new molecules. The cause of nutrition and growth lies not in the organism as a whole, but in the separate elementary parts–the cells. That in point of fact every cell, when separated from the organism, is not capable of further growth as little militates against this theory, as its incapability of existing separate from the swarm would be an argument against the independent life of a bee...... The inquiry into the fundamental forces of organisms, therefore, is reduced into one concerning the fundamental forces of the single cells." (p. 229.)
And yet, strongly as Schwann maintains, not only here but in many other places, the view that the vital forces are manifested by the cells as machines, and are not inherent in the matter of which these cells are composed, apart from their form; he gives it up in effect when he comes to treat of these forces in detail. The fundamental cell-forces are, he says, of two kinds, the attractive and the metabolic, the former regulating growth, the latter determining the chemical changes; and he shows very justly that these forces are not located in any special centres in the cell, but are exhibited by all its solid constituents (pp. [262] 233, 236), and that they may be exhibited by different portions of these solid constituents, and to a different extent by these different portions (p. 233); proving hereby, very clearly, as it seems to us, that the forces in question are not centralized in the cells, but are resident in their component molecules. All Schwann's able comparison of cell-development with crystallization tends, in fact, to the same conclusion. When matter crystallizes from a solution, the presence of a foreign body may determine the place and form of the deposit; but the crystals themselves are the result, not of the attractive forces of the foreign body, but of the forces resident in their component molecules. So in cell-development, if it is to be rigorously compared to crystallization, even if the nuclei represent the foreign bodies, which determine the place of the chemical and morphological alterations in the surrounding substance, it by no means follows that they are their cause.
Kölliker (§§ 11-13), resting especially upon the phenomena of yelk-division and of endogenous cell-development, advocates the existence of a peculiar molecular attraction proceeding from the nucleolus first, and subsequently from the nucleus. Now as regards endogenous cell-development, we must confess that we can find no more ground for its occurrence among animals than among plants. Nägeli's cell-development around portions of contents, upon which Kölliker lays so much stress, is nothing more than a case of division of the endoplast (primordial utricle) and subsequent development of periplastic substance round the portions. In cartilage, which is so often quoted as offering marked endogenous cell-development, we must agree with Leidy and Remak, that nothing but division of the endoplasts (nuclei, primordial utricles) and ingrowth of the periplast (intercellular substance, cell-wall) occurs. In these endoplasts again, the very existence of a nucleus is in the highest degree variable and inconstant, and division occurs as well without it as with it.
The process of yelk-division–that remarkable manifestation of a tendency to break up, in the yelk of most animals, into successively smaller spheroids, in each of which a nucleus of some kind appears–seems, at first, to offer very strong evidence in favour of the exertion of some attraction by these nuclei upon the vitelline mass. But we think that a closer examination completely deprives this evidence of all weight. In the first place, the appearance of the nuclei is in many cases subsequent to segmentation. It is thus in Strongylus auricularis (Reichert), in Phallusia (Krohn), in the hen's egg (Remak). In the second place, it seems difficult to conceive any mode of operation of a central attractive force which shall give rise to the phenomena of segmentation, for the resulting spheroids always pass into one another [263] by extensive plane surfaces, whereas the even action of two attractive centres, in a mass free to move, would give rise to two spheroids in contact only by a point. Again, Remak has observed that in the frog's egg the time occupied by the formation of the groove, indicating the first line of cleavage upon the upper half of the yelk, is very much shorter than that required to give rise to the corresponding line upon the lower half–a fact which is quite unintelligible upon the theory of a central attraction.
Thirdly, in Cucullanus, Ascaris dentata, &c., Kölliker has shown that, though nuclei are developed, no yelk-division occurs; and in the later stages of division of the frog's egg yelk masses are found undivided, and containing many nuclei.
Finally, in Ascaris mystax, according to Dr. Nelson,13 the embryonic vesicles absolutely revolve in circles during the progress of yelk-division–-a phenomenon which seems incompatible with the existence of any mutual attractive reaction between themselves and the vitelline mass.
We see, in short, that the effects of the force supposed to be exerted by the nuclei may take place without them, and, on the other hand, that the nuclei may be present without exerting the peculiar forces which they are supposed to possess; and finally that even if such forces exist, they must be something very different from all the attractive forces of which we have any conception; and therefore that the hypothesis of nuclear force is no explanation, but merely a fresh name for the difficulty.
We are as little able to discover any evidence of the existence of metabolic forces in the nuclei. The metabolic changes of the tissues–such as we see, for instance, in the conversion of cartilage into bone, of cartilage into connective tissue–do not take place, either primarily or with greater intensity, in the neighbourhood of the nuclei; a fact of which striking evidence is afforded by ossifying cartilage, in which the first deposit of calcareous matter occurs, not in areas surrounding each nucleus, as we should expect if they exerted a metabolic influence, but in straight lines, which stretch from the ossified surface into the substance of the matrix of the cartilage, and the amount of calcareous matter in which gradually diminishes as we recede from the ossified part, without the least reference to the nuclei. It is the same with the metamorphosis of the periplast of cartilage when it passes into tendon.
From all this we consider it to be satisfactorily shown, that there is no evidence that the "cells" of living bodies are, in any respect, centres [264] of those properties, which are called vital forces. What, then, are these cells? it may be asked;–what is the meaning of the unquestionable fact that the first indication of vitality, in the higher organisms at any rate, is the assumption of the cellular structure?
In answering these questions, we would first draw attention to the definition of the nature of development in general, first clearly enunciated by Von Baer. "The history of development," he says, "is the history of a gradually increasing differentiation of that which was at first homogeneous." The yelk is homogeneous; the blastoderma is a portion of it which becomes different from the rest, as the result of the operation of the laws of growth; the blastoderma, again, comparatively homogeneous, becomes differentiated into two or more layers; the layers, originally identical throughout, set up different actions in their various parts, and are differentiated into dorsal and visceral plates, chorda dorsalis and bodies of vertebræ, &c., &c. No one, however, imagines that there is any causal connexion between these successive morphological states. No one has dreamt of explaining the development of the dorsal and visceral plates by blastodermic force, nor that of the vertebræ by chorda-dorsalic force. On the other hand, all these states are considered, and justly, to result from the operation of some common determining power, apart from them all–to be, in fact, the modes of manifestation of that power.
Now, why should we not extend this view to histology, which, as we have explained, is only ultimate morphology? As the whole animal is the result of the differentiation of a structureless yelk, so is every tissue the result of the differentiation of a structureless blastema–the first step in that differentiation being the separation of the blastema into endoplast and periplast, or the formation of what is called a "nucleated cell."14 Then, just as in the development of the embryo, when the blastodermic membrane is once formed, new organs are not developed in other parts of the yelk, but proceed wholly from the differentiation of the blastoderm,–so histologically, the "nucleated cell," the periplast with its endoplast, once formed, further development takes place by their growth and differentiation into new endoplasts and periplasts. The further change into a special tissue, of course, succeeds and results from this primary differentiation, as we have seen the bodies of the vertebræ succeed the chorda dorsalis; but is there any more reason for supposing a causal connexion between the one pair of phenomena than between the other? The cellular structure precedes the special structure; but is the latter, therefore, the result of [265] a "cell-force," of whose existence there is on other grounds no evidence whatever? We must answer in the negative. For us the primarily cellular structure of plants and animals is simply a fact in the history of their histological development–a histologically necessary stage, if one may so call it, which has no more causal connexion with that which follows it, than the equally puzzling morphological necessity for the existence of a chorda dorsalis or of Wolffian bodies has with the development of the true vertebræ or of the true kidneys.
If this be true, we might expect, as we find, that the differentiation of the germinal disc, for instance, into a primitive groove and lateral portions–the first stage of development in the embryo of all vertebrate animals–does not occur in mollusks; as we find, again, that the differentiation of the embryo into plumula and cotyledons which occurs in a great number of plants is absent in others; so if, like these, the histological differentiation into cells have no necessary causal connexion with the action of the vital forces, but be merely a genetic states we may expect to meet with cases in which it does not occur. Such, in fact, are the so-called unicellular plants and animals–organisms which often exhibit no small complexity of external form; but present no internal histological differentiation. In the genus Caulerpa we have an Alga presenting apparent leaves, stems, and roots, and yet which, according to Nägeli, consists of a single cell–that is, is not composed of cells at all. The Vorticellæ furnish us with examples of animals provided with a distinct œsophagus, a muscular pedicle, &c., and yet in which no further histological differentiation can be made out. As Wolff15 says–
"The latter (Roesel's Proteus) has no structure, no determinate figure, and even the indeterminate figure that it has at any given time does not remain the same, but alters continually. We can, in fact, regard all these plants and animals as little else than living or vegetating matters–hardly as organized bodies.
"§ 74. However, all these plants and animals nourish themselves, vegetate, and propagate their species, just as well and as easily as the most artificial pieces of mechanism to be met with in the vegetable or animal kingdom."
It is true, indeed, that the difficulty with regard to these organisms has been evaded by calling them "unicellular"–by supposing them to be merely enlarged and modified simple cells; but does not the phrase an "unicellular organism" involve a contradiction for the cell-theory? In the terms of the cell-theory, is not the cell supposed to be an [266] anatomical and physiological unity, capable of performing one function only–the life of the organism being the life of the separate cells of which it is composed? and is not a cell with different organs and functions something totally different from what we mean by a cell among the higher animals? We must say that the admission of the existence of unicellular organisms appears to us to be virtually giving up the cell-theory for these organisms. If it be once admitted that a particle of vitalizable matter may assume a definite and complex forms may take on different functions in its different parts, and may exhibit all the phenomena of life, without assuming the cellular structure, we think that it necessarily follows that the cells are not the centres of the manifestation of the vital forces; or that, if they be so, the nature of these forces is different in the lower organisms from what it is in the higher–a proposition which probably few would feel disposed to maintain.
So much for the critical, and therefore more or less ungrateful, portion of our task. We have seen how the great idea, fully possessed by Fallopius, that life is not the effect of organization, nor necessarily dependent upon it, but, on the other hand, that organization is only one of the phenomena presented by living matter–carried to absurdity by Stahl and Van Helmont–has, on the other hand, been too much neglected by the later writers who have attempted to reduce life to the mere attractions and repulsions of organic centres, or to consider physiology simply as a complex branch of mere physics. We have seen how this latter notion has been fostered by the misconceptions of a great botanist, only too faithfully followed in the animal world by the illustrious author of the cell-theory; and we have endeavoured to show how the solitary genius of Wolff had kept in the old track, and that the choice of modern histologists lies between him and Schleiden and Schwann. It will be sufficiently obvious that our own election has long been made in this matter, and we beg to submit the following sketch of a general theory of the structure of plants and animals–conceived in the spirit, and not unfrequently borrowing the phraseology, of Wolff and Von Baer.
Vitality, the faculty, that is, of exhibiting definite cycles of change in form and composition, is a property inherent in certain kinds of matter.
There is a condition of all kinds of living matter in which it is an amorphous germ–that is, in which its external form depends merely on ordinary physical laws, and in which it possesses no internal structure.
Now, according to the nature of certain previous conditions–the [267] character of the changes undergone–of the different states necessarily exhibited–or, in other words, the successive differentiations of the amorphous mass will be different.
Conceived as a whole, from their commencement to their termination, they constitute the individuality of the living being, and the passage of the living being through these states is called its development. Development, therefore, and life are, strictly speaking, one thing, though we are accustomed to limit the former to the progressive half of life merely, and to speak of the retrogressive half as decay, considering an imaginary resting point between the two as the adult or perfect state.16
The individuality of a living thing, then, or a single life, is a continuous development, and development is the continual differentiation, the constant cyclical change of that which was, at first, morphologically and chemically indifferent and homogeneous.
The morphological differentiation may be of two kinds. In the lowest animals and plants–the so-called unicellular organisms–it may be said to be external, the changes of form being essentially confined to the outward shape of the germ, and being unaccompanied by the development of any internal structure.
But in all other animals and plants, an internal morphological differentiation precedes or accompanies the external, and the homogeneous germ becomes separated into a certain central portion, which we have called the endoplast, and a peripheral portion, the periplast. Inasmuch as the separate existence of the former necessarily implies a cavity, in which it lies, the germ in this state constitutes a vesicle with a central particle, or a "nucleated cell."
There is no evidence whatever that the molecular forces of the living matter (the "vis essentialis" of Wolff, or the vital forces of the moderns) are by this act of differentiation localized in the endoplast, to the exclusion of the periplast, or vice versa. Neither is there any evidence that any attraction or other influence is exercised by the one over the other; the changes which each subsequently undergoes, though they are in harmony, having no causal connexion with one another, but each proceeding, as it would seem, in accordance with the general determining laws of the organism. On the other hand, the "vis essentialis" appears to have essentially different and independent ends in view–if we may for the nonce speak metaphorically–in thus separating the endoplast from the periplast.
[268] The endoplast grows and divides; but, except in a few more or less doubtful cases, it would seem to undergo no other morphological change. It frequently disappears altogether; but, as a rule, it undergoes neither chemical nor morphological metamorphosis. So far from being the centre of activity of the vital actions, it would appear much rather to be the less important histological element.
The periplast, on the other hand, which has hitherto passed under the names of cell-wall, contents, and intercellular substance, is the subject of all the most important metamorphic processes, whether morphological or chemical, in the animal and in the plant. By its differentiation, every variety of tissue is produced; and this differentiation is the result not of any metabolic action of the endoplast, which has frequently disappeared before the metamorphosis begins, but of intimate molecular changes in its substance, which take place under the guidance of the "vis essentialis," or, to use a strictly positive phrase, occur in a definite order, we know not why.
The metamorphoses of the periplastic substance are twofold–chemical and structural. The former may be of the nature either of conversion: change of cellulose into xylogen, intercellular substance, &c., of the indifferent tissue of embryos into collagen, chondrin, &c.; or of deposit: as of silica in plants, of calcareous salts in animals.
The structural metamorphoses, again, are of two kinds–vacuolation, or the formation of cavities; as in the intercellular passages of plants, the first vascular canals of animals; and fibrillation, or the development of a tendency to break up in certain definite lines rather than in others, a peculiar modification of the cohesive forces of the tissue, such as we have in connective tissue, in muscle, and in the "secondary deposits" of the vegetable cell.
Now to illustrate and explain these views, let us return to the vegetable and animal tissues, as we left them in describing the base of the Sphagnum leaf and fœtal cartilage, and trace out the modification of these, which are identical with all young tissues, into some of the typical adult forms.
The point of the Sphagnum leaf is older than the base, and it is easy to trace every stage from the youngest to the complete forms in this direction. At the base of the leaf we find, as has been said, nothing but minute endoplasts, each resembling the other, embedded in a homogeneous periplastic substance (A); as we trace these upwards, we find that some of the endoplasts increase in size more rapidly than the others (B), and eventually totally disappear, leaving only the endoplastic cavity, or "cell," which contained them. In the surrounding cells, the endoplasts are very obvious as granular primordial utricles [269] (C).
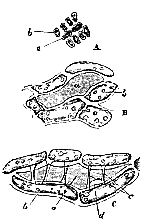
Fig. 2. Portions of the leaf of Spagnum. A, from the base; B, more towards the point; C, fully formed. a, endoplasts which disappear; b, those which remain; c, spiral thickenings of periplast in the cavities of the former; d, apertures formed by resorption.
After the disappearance of the endoplast, changes commence in the periplastic substance or wall of the cell (a), more or less circular or spiral thickenings (c) taking place in it, so as to form the well-known fibre-cell of the sphagnum leaf; and at the same time, a process of resorption occurs in particular parts of the wall, so that round apertures are formed (d). Nothing can be more instructive than this case, the leaf being composed of a single layer of delicate and transparent cells, so that there are no interfering difficulties of observation; and we see demonstrated, in the most striking manner, that the endoplast or primordial utricle has nothing to do with the metamorphoses which occur in the periplastic substance. The disappearance of the primordial utricle in cells which are undergoing thickening was, in truth, long ago pointed out by Von Mohl; but neither he nor any of his successors seem to have noticed how completely this fact does away with that activity of the primordial utricle, and passivity of the cell-wall, which they all assume. We have here, in fact, the cell-wall commencing and carrying through its morphological changes after the primordial utricle has completely disappeared, and we see that the so-called secondary deposit in this case is a morphological differentiation of the periplast, which at the same time exhibits its peculiar powers by setting up a resorption of its substance at another point. Here however, we have no marked chemical differentiation; for an instance of which we may turn to the collenchyma of the beet-root (fig. 1, A). There is no question that, at one period of its development, the whole periplastic substance here, as in the Sphagnum, was homogeneous, and of the same chemical constitution. In the fully formed beet-root, however, we have no less than three compounds disposed around each cell cavity. The periplastic substance has, in fact, undergone both a chemical and a morphological differentiation–the innermost layers (c) consisting of ordinary cellulose; [270] the next of a substance which swells up in water (b); and the outermost of a different, but not exactly defined, substance (a). We may call one of these portions "cell-membrane," and another intercellular substance, but they are assuredly all nothing but differentiated portions of one and the same periplast.
Woody tissue presents precisely the same phenomena, the inner layers of the periplastic substance having, very generally, a different composition from the outer.
Morphologically, we have already noticed the lamination of the periplastic substance, and we may mention its fibrillation, a process which takes place almost invariably in the inner layers of the periplast, and to which the well-known spirality of the so-called secondary deposits must be referred; but a more important process for our present purpose is what we have called Vacuolation; the development of cavities in the periplastic substance independent of the endoplast, and which, to distinguish them from the cells, may conveniently be termed Vacuolæ. In the youngest vegetable tissues there are no such cavities, the periplastic substance forming a continuous solid whole; and it is by this vacuolation, which occurs as the part grows older, that all the intercellular passages are formed, and that many cells obtain that spurious anatomical independence to which we have adverted above. The exaggerated development of the vacuolæ in the pith of the rush converts the periplastic substance, with its proper endoplastic cavities, into regular stellate cells. (Fig. 1, B.)
Sufficient has been said to illustrate the differentiation of the primitive vegetable structure into its most complex forms. If we turn to the animal tissues, we shall find the same simple principles amply sufficient to account for all their varieties.
In the plant, as we have seen, there are but two histological elements–the periplastic substance and the endoplasts, cell-wall and intercellular substance, being merely names for differentiated portions of the former; cell-contents, on the other hand, representing a part of the latter. In the animal, on the other hand, if we are to put faith in the present nomenclature, we find cell-wall, intercellular substance, and cell-contents, forming primitive elements of the tissues, and entering into their composition as such: there have been no small disputes whether the collagenous portion of connective tissue is intercellular substance or cell-wall, the elastic element being pretty generally admitted to be developed from distinct cells. Again, it appears to be usual to consider the fibrillæ of striped muscle as modified cell-contents, while the sarcolemma represents the cell-walls. The hyaline substance of cartilage is asserted by some to be cell-wall, by some to be [271] intercellular substance; while the walls of the epithelium cavities are admitted on all hands to be cell-walls. We confess ourselves quite unable to find any guiding principle for this nomenclature, unless it be that the toughest structure surrounding a "nucleus" is to be taken as cell-wall, anything soft inside it being contents, and anything external to it intercellular substance; which is hardly a caricature of the vagueness which pervades histological works upon this subject. The results, we think, from the attempt to determine the homology of the parts of the tissues having been made from the examination of their embryonic conditions, where it is often very obscure, and hardly to be made out. It is another matter if we adopt the "principle of continuity" of Reichert–a method of investigation which has been much neglected. This principle is simply, that whatever histological elements pass into one another by insensible gradations are homologous and of the same nature; and it is so clear and easy of application, that we can but wonder at its hitherto limited use. We will now proceed to analyze the nature of the constituents of some of the most characteristic tissues in this way, starting from that of embryonic cartilage, as we have described it above.
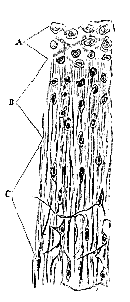
Fig. 3. Junction of tendo-Achillis and cartilage of the calcaneum in a kitten. A, Pure cartilage; B, intermediate portion; C, tendon. It must be understood that the transition is in reality much more gradual, the different stages having here been approximated for the sake of economizing space.
Connective tissue occurs in two forms,–which, however, pass into one another by infinite gradations,–the solid and the areolated: of the former we may take a tendon as an example; of the latter, the loose areolar tissue, which is found forming the inner layer of the skin and mucous membranes. Fig. 3 represents the junction between the tendo-Achillis and the cartilage of the os calcis in a young kitten. At A we have pure cartilage, the endoplasts lying within cavities whose walls present more or less defined contours. At B, the cavities and their contained endoplasts are somewhat elongated, and a faint striation is obvious in the upper portion of the periplastic substance, which [272] becomes stronger and stronger as we proceed lower down, until it ends in an apparent fibrillation. A chemical change has at the same time taken place, so that in this portion the striated part of the periplast is swollen up more or less by acetic acid, the walls of the cavities remaining unaffected, and thence becoming more distinct; while in the portion A, the whole periplast was nearly equally insensible to this re-agent. The portion C, nearest the tendon, and passing into it, is completely tendinous in its structure. The periplast exhibits strong fibrillation, and is very sensitive to acetic acid, while not only the walls of the cavities, but the intermediate periplast, in certain directions, which radiate irregularly from them, have changed into a substance which resists acetic acid even more than before, and is in fact elastic tissue. Compare this process with that which we have seen to be undergone by the collenchyma of the beetroot, and we have the fibrillation of the outer portion of the periplast around each endoplast, and its conversion into collagen, answering to the lamination of the "intercellular substance," and its conversion into a vegetable gelatinous matter, while the elastic corresponds with the cellulose inner wall.
Fig. 4. Stellate "cells" of young connective tissue from the actinenchyma of the enamel organ of the calf.
The testimony of numerous observers agrees that cartilage is converted into connective tissue in the way described. Professor Kölliker, who unwillingly admits the fact, suggests, nevertheless, that such connective tissue, inasmuch as it presents differences in its mode of development, the collagenous element in the latter being always developed from cells.17
Now, we might be inclined to ask, if the substance of the tendo-Achillis is not connective tissue, but only "täuschend ähnlich," what is? But it is better to attack Prof. Kölliker's stronghold, the areolated [273] gelatinous connective tissue, which is, as he justly observes, the early form of fœtal connective tissue generally. (l. c., p. 58.) If the outer layer of the corium of the skin, or the submucous gelatinous tissue in the enamel organ, be teased out with needles, we shall obtain various stellate or ramified bodies containing endoplasts (fig. 4), which Kölliker calls cells, and which, as he states, do assuredly pass into and become bundles of fibrillated connective tissue. But is this really a different mode of development from that already described? We think not. Indeed if that portion of this young gelatinous connective tissue, which lies immediately adjacent to the epidermis or epithelium, be examined, it will be found to present a structure in all respects similar to fœtal cartilage, that is, there is a homogeneous matrix in which the endoplasts are dispersed (fig. 5, B). If this be traced inwards, it will be found that the endoplasts become more widely separated from one another, and that the matrix in places between them is softened and altered, while in their immediate neighbourhood, and in the direction of irregular lines stretching from them, it is unaltered.
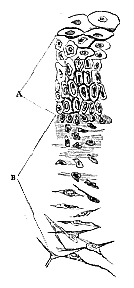
Fig. 5. Submucous tissue and epithelium of the tongue of the kitten. A, Epithelium; B, young connective tissue
This is, in fact, the first stage of that process which we have called vacuolation. In this condition the intermediate softened spots still retain sufficient consistence not to flow out of a section; but yielding, as it does, in these localities, much more readily than in others, it is easy enough to tear out the firmer portion in the shape of "cells," which are fusiform, irregular, or stellate; and the whole tissue has therefore been described (Reichert, Virchow, Schwann) as consisting of cells connected by an "intercellular substance." Both "cell-walls and "intercellular substance," however, are portions of the same periplast, and together correspond with the matrix of the cartilage. When, therefore, in the course of further development, the "intercellular substance" becomes quite fluid and so disappears, the outer portion of these cells being converted into fibrillated collagenous tissue, and the inner into elastic substance, we have, notwithstanding the apparently great difference [274] in reality exactly the same mode of metamorphosis of the same elements as in the preceding instance. Connective tissue, therefore, we may say, consists in its earliest state of a homogeneous periplast inclosing endoplasts. The endoplasts may elongate to some extent, but eventually become lost, and cease, more or less completely, to be distinguishable elements of the tissue. The periplast may undergo three distinct varieties of chemical differentiation, e.g., into the gelatinous "intercellular substance," the collagenous "cell-wall," and the elastic "cell-wall;" and two varieties of morphological differentiation, vacuolation, and fibrillation–and the mode in which these changes take place gives rise to the notion that the perfect tissue is composed of elements chemically and mechanically distinct.
The proper understanding of the nature and mode of development of the component parts of connective tissue is, we believe, of the first importance in comprehending the other tissues. If we clearly bear in mind, in the first place, that the periplast is capable of undergoing modifications quite independently of the endoplasts; and, secondly, that in consequence of their modification, elements may become optically, mechanically, or chemically separable from a perfect tissue which were not discoverable in its young form, and never had any separate existence; many of the great difficulties and perplexities of the cell-theory will disappear. Thus, for instance, with regard to the structure of bone, there can be no doubt that the "nuclei" of the corpuscles are endoplasts, and that the calcified matrix is the periplast. This calcified matrix has, however, in adult bone, very often a very regular structure, being composed of definite particles. To account for these, Messrs. Tomes and De Morgan, in their valuable essay on ossification, which has just appeared,18 suppose that certain "osteal cells" exist and become ossified. We have no intention here of entering upon the question of the existence of these "osteal cells" as a matter of fact, but we may remark that they are by no means necessary, as the appearance might arise from a differentiation of the periplast into definite particles, corresponding with that which gives to connective tissue its definite and fibrillated aspect. So with regard to the vexed question whether the lacunæ have separate parietes or not, how readily comprehensible the opposite results at which different observers have arrived become, if we consider that their demonstrability or otherwise results simply from the nature and amount of the chemical difference which has been established in the periplast in the immediate neighbourhood of the endoplast with regard to that in the rest of the periplast. In fig. 3 substitute calcific for collagenous metamorphosis, [275] and we should have a piece of bone exhibiting every variety of lacunæ, from those without distinct walls to those which constitute regular stellate "bone corpuscles." Finally, in bone, the formation of the "Haversian spaces" of Tomes and De Morgan is a process of vacuolation, strictly comparable to that which we have described as giving rise to the areolated connective tissue. Cancellated bone is, in fact, areolated osseous tissue. Once having comprehended the fact that the periplast is the metamorphic element of the tissues, and that the endoplast has no influence nor importance in histological metamorphosis, there ceases to be any difficulty in understanding and admitting the development of the tubules of the dentine and the prisms of the enamel, without the intervention of endoplasts. These are but the extreme and obvious cases in which nature has separated for us two histological elements and two processes, which are elsewhere confounded together.

Fig. 6. Continuity of muscle with connective tissue from the tongue of the lamb. A, Connective tissue; B, muscle.
One of the most complicated of tissues is striped muscle, yet the true homology of its elements seems to us to become intelligible enough upon these principles. Dr. Hyde Salter has pointed out,19 that in the tongue the muscles pass directly into the bundles of the submucous connective tissue which serve as their tendons. We have figured such a transition in fig. 6. The tendon A may be seen passing insensibly into the muscle B, the granular sarcous elements of the latter appearing as it were to be deposited in the substance of the tendon (just as the calcareous particles are deposited in bone), at first leaving the tissue about the walls of the cavities of the endoplasts, and that in some other dimensions unaltered. These portions, which would have represented the elastic element in ordinary connective tissue, disappear in the centre of the muscular bundle, and the endoplasts are immediately surrounded by muscle, just as, in many specimens of bone, the lacunæ have no distinguishable walls. On the other hand, at the surface of the bundle the representative of the elastic element remains, and often becomes much developed as the sarcolemma. There is no question here of muscle resulting from the [276] contents of fused cells, &c. It is obviously and readily seen to be nothing but a metamorphosis of the periplastic substance, in all respects comparable to that which occurs in ossification, or in the development of tendon. In this case we might expect that as there is an areolar form of connective tissue, so we should find some similar arrangement of muscle; and such may indeed be seen very beautifully in the terminations of the branched muscles, as they are called. In fig. 7 the termination of such a muscle, from the lip of the rat, is shown, and the stellate "cells" of areolated connective tissue are seen passing into the divided extremities of the muscular bundle, becoming gradually striated as they do so.
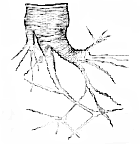 Fig. 7. Branched muscle, ending in stellate connective "cells," from the upper lip of the rat.
Fig. 7. Branched muscle, ending in stellate connective "cells," from the upper lip of the rat.
We have already exceeded our due limits, and we must therefore reserve for another place the application of these views to other tissues. There is, however, one application of the mode of termination of the branched muscles to which we have just referred, which is of too great physiological importance to be passed over in silence. In the muscle it is obvious enough that whatever homology there may be between the stellate "cells" and the muscular bundles with which they are continuous, there is no functional analogy, the stellate bodies having no contractile faculty. But a nervous tubule is developed in essentially the same manner as a muscular fasciculus, the only difference being that fatty matters take the place of syntonin. Now, it commonly happens that the nerve-tubules terminate in stellate bodies of a precisely similar nature; and these, in this case, are supposed to possess important nervous functions, and go by the name of "ganglionic cells." From what has been said, however, it is clear that these may be genetically and not functionally connected with the nervous tubules, and that, so far from being the essential element of the nervous centres and expansions, it is possible that the "ganglionic cells" have as little nervous function as the stellate cells in the lip of the rat have contractile function.
[277] We cannot conclude better than by concisely repeating the points to which we have attempted to draw attention in the course of the present article.
We have endeavoured to show that life, so far as it is manifested by structure, is for us nothing but a succession of certain morphological and chemical phenomena in a definite cycle, of whose cause or causes we know nothing; and that, in virtue of their invariable passage through these successive states, living beings have a development, a knowledge of which is necessary to any complete understanding of them. It has been seen that Von Baer enunciated the law of this development, so far as the organs are concerned; that it is a continually increasing differentiation of that which was at first homogeneous; and that Caspar Friedrich Wolff demonstrated the nature of histological development to be essentially the same, though he erred in some points of detail, we have found Schwann demonstrating for the animal, what was already known for the plant–that the first histological differentiation, in the embryo, is into endoplast and periplast, or, in his own phrase, into a "nucleated cell;" and we have endeavoured to show in what way he was misled into a fundamentally erroneous conception of the homologies of these two primitive constituents in plants and animals–that what he calls the "nucleus" in the animal is not the homologue of the "nucleus" in the plant, but of the primordial utricle.
We have brought forward evidence to the effect that this primary differentiation is not a necessary preliminary to further organization–that the cells are not machines by which alone further development can take place, nor, even with Dr. Carpenter's restriction (p. 737), are to be considered as "instrumental" to that development. We have tried to show that they are not instruments, but indications–that they are no more the producers of the vital phenomena than the shells scattered in orderly lines along the sea-beach are the instruments by which the gravitative force of the moon acts upon the ocean. Like these, the cells mark only where the vital tides have been, and how they have acted.
Again, we have failed to discover any satisfactory evidence that the endoplast, once formed, exercises any attractive, metamorphic, or metabolic force upon the periplast; and we have therefore maintained the broad doctrine established by Wolff, that the vital phenomena are not necessarily preceded by organization, nor are in any way the result or effect of formed parts, but that the faculty of manifesting them resides in the matter of which living bodies are composed, as such–[278] or, to use the language of the day, that the "vital forces" are molecular forces.
It will doubtless be said by many, But what guides these molecular forces? Some Cause, some Force, must rule the atoms and determine their arrangement into cells and organs; there must be something, call it what you will–Archæus, "Bildungs-trieb," "Vis Essentialis," Vital Force, Cell-force–by whose energy the vital phenomena in each case are what they are.
We have but one answer to such inquiries: Physiology and Ontology are two sciences which cannot be too carefully kept apart; there may be such entities as causes, powers, and forces, but they are the subjects of the latter, and not of the former science, in which their assumption has hitherto been a mere gaudy cloak for ignorance. For us, physiology is but a branch of the humble philosophy of facts; and when it has ascertained the phenomena presented by living beings and their order, its powers are exhausted. If cause, power, and force mean anything but convenient names for the mode of association of facts, physiology is powerless to reach them. It is satisfactory to reflect, however, that in this comparatively limited sphere the inquiring mind may yet find much occupation.
1 It is amusing to find M. Comte, a mere bookman on these subjects, devoting a long argument (Philosophie Positive, tom. iii. p. 288) to a refutation (?) of what he calls the "profonde irrationalité" of Bichât's definition. As a specimen of the said refutation, we may select the following passage: "Si comme le supposait Bichât, tout ce qui entoure les corps vivans tendait réellement à les detruire, leur existence serait par cela même radicalement inintelligible; car, où pourraient-ils puiser la force nécessaire pour surmonter même temporairement un tel obstacle?" What a question for a positive philosopher! Does M. Comte doubt his own power to get up from his easy chair, because it is unquestionably true that the action of the whole globe "tends" to retain him in his sitting posture, and because he cannot tell whence he gets the force which enables him to rise? 2 It is not easy to frame a definition of the differences between living and not living bodies which shall perfectly defy cavil. That in the text–based on the inertia of not living bodies, the internal activity of living bodies–marks the difference strongly but not unobjectionably, for it might be said that a nebula undergoing change would, by this definition, be a living body, and in the next place, it might be urged how do we know that the activity of living bodies is not really the result of some external cause with which we are unacquainted? It might be said, that the apparent absence of change in external conditions, is no more evidence that the vital phenomena are independent of some such causes, than the continuous running of a stream when the dam is opened, independent of any further alteration of external conditions, is evidence of spontaneity. The action of the spermatozoon, e.g., might be compared to the raising of the dam. We have preferred above, however, a vivid to an exact definition of our conception of life, as likely to be more useful. If we were to attempt an exact definition, it would be, that a living being is a natural body which presents phenomena of growth, of change of form, and of chemical composition, of a definite nature, and occurring in definite cycles of succession. This definition will separate living beings from all other terrestrial bodies. It separates them from cosmical bodies (nebulæ, &c.) only by the nature of the phenomena which succeed one another; so true is it that the microcosm and the macrocosm are reflections of one another. 3 Wolff: Theorie von der Generation, p. 12. 4 Terms by no means always convertible, but which may for the present be taken to be so. 5 Compare Kölliker's Handbuch, Introduction; or Sprengel's Geschichte der Arzneykunde. 6 Histoire des Sciences Naturelles. 7 The world, always too happy to join in toadying the rich, and taking away the "one ewe lamb" from the poor, persists in ascribing the theory of the metamorphosis of plants to Göthe, in spite of the great poet himself (see Göthe's Werke, Cotta, 1840, B. 36, p. 105: "Entdeckung eines trefflichen Vorarbeiters "), who not only acknowledges his own obligations to Wolff, but speaks with just wonder and admiration of the 'Theoria Generationis,' the work of a young man of six-and-twenty. 8 Theorie von der Generation, § 64. 9 Theoria Generationis, and Von der eigenthümlichen Kraft, p. 48. 10
It is very curious to find even Schwann's definition of cell-development as the "crystallization of a permeable body" anticipated by Wolff, Von d. eigenthumlichen Kraft, &c., p. 63: "......so, from the case of many other attractions, especially of crystallization, which, among all known phenomena, comes nearest to vegetation, that without the second property–viz., that by means of which the mutually attractive substances interpenetrate and mingle with one another–the attractive force, although it should possess the first property, yet could as little effect nutrition. The particles of a salt dissolved in water attract only particles of salt–i.e., homogeneous substances, and repel all heterogeneous matters; for we get pure (crystals of the) salts. But out of this attraction comes nothing that can be compared with nutrition; for although the whole mass of substance is increased by degrees, yet crystals once formed remain as they are, and are not increased in their substance, and nothing less than the formation of new organization and continual change of figure accompanies this increase....... Crystals once formed attract the saline particles only to their outer surface, upon which the attracted parts are deposited. They do not attract these particles into their substance ......If the saline particles, on the other hand, penetrated the crystals and increased their substance homogeneously in all parts, this process would be indistinguishable from nutrition, and would consequently be a true nutrition." Compare Schwann, pp. 239–257. 11 On Alex. Braun's authority, Ueber Verjüngung, &c., p 186. 12 Henfrey, Linn. Trans. 1853. 13 Philosophical Transactions, 1852. 14 Compare Reichert, p. 35. 15 'Von der wesentlichen Kraft,’ p. 40. 16 Dr. Lyons, in his interesting 'Researches on the Primary Stages of Histogenesis and Histolysis,' has invented a most convenient and appropriate term for this latter half of development, so far as the tissues are concerned–viz., Histolysis. 17 Handbuch, pp. 58, 59, 218. 18 Phil. Trans., 1853. 19 Art. 'Tongue,' Todd's Cyclopædia.
|
THE
HUXLEY
FILE

|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
{kind=link}