[538] The necessity of discussing so great a subject as the Theory of the Vertebrate Skull in the small space of time allotted by custom to a lecture, has its advantages as well as its drawbacks. As, on the present occasion, I shall suffer greatly from the disadvantages of the limitation, I will, with your permission, avail myself to the uttermost of its benefits. It will be necessary for me to assume much that I would rather demonstrate, to suppose known much that I would rather set forth and explain at length; but on the other hand, I may consider myself excused from entering largely either into the history of the subject, or into lengthy and controversial criticisms upon the views which are, or have been, held by others.
The biological science of the last half-century is honorably distinguished from that of preceding epochs, by the constantly increasing prominence of the idea, that a community of plan is discernible amidst the manifold diversities of organic structure. That there is nothing really aberrant in nature; that the most widely different organisms are connected by a hidden bond; that an apparently new and isolated structure will prove, when its characters are thoroughly sifted, to be only a modification of something which existed before,–are propositions which are gradually assuming the position of articles of faith in the mind of the investigators of animated nature, and are directly, or by implication, admitted among the axioms of natural history.
And this is not wonderful; for no living being can be attentively [539] studied without bearing witness to the truth of these propositions. The tyro in comparative anatomy cannot fail to be struck with resemblances between the leg and the jaw of a crustacean; between the parts of the mouth of a beetle and those of a bee; between the wing of the bird and the fore-limb of the mammal. Everywhere he finds unity of plan, diversity of execution.
Or again, how can the intelligent student of the human frame consider the backbone, with its numerous joints or vertebræ, and trace the gradual modification which these undergo downwards into the sacrum and coccyx, and upwards into the atlas and axis, without the notion of a vertebra in the abstract, as it were, gradually dawning upon his mind; the conception of an ideal something which shall be a sort of mean between these various actual forms, each of which may then easily be conceived as a modification of the abstract or typical vertebra?
Such an idea, once clearly apprehended, will hardly permit the mind which it informs to rest at this point. A glance at a section of that complex bony box formed by the human skull and face shows that it consists of a strong central mass, whence spring an upper arch and a lower arch. The upper arch is formed by the walls of the cavity containing the brain, and stands in the same relation to it, as does the neural arch of a vertebra to the spinal cord, with which that brain is continuous. The lower arch encloses the other viscera of the head, in the same way as the ribs embrace those of the thorax. And not only is the general analogy between the two manifest but a young skull may be readily separated into a number of segments, in each of which it requires but little imagination to trace a sort of family likeness to such an expanded vertebra as the atlas.
What can be more natural then than to take another step–to conceive the skull as a portion of the vertebral column still more altered than the sacrum or the coccyx, whose vertebræ are modified in correspondence with the expansion of the anterior end of the nervous centre and the needs of the cephalic end of the body, just as those of the sacrum are fashioned in accordance with the contraction of the nervous centre and the mechanical necessities of the opposite extremity of the frame?
Two generations have passed away since, perhaps, by some such train of reasoning as this, such a conception of the nature of the vertebrate skull arose in the mind of the philosophic poet, Goethe; and a somewhat shorter period has elapsed since a poetical, or perhaps I might more justly say a fanciful, philosopher, Oken, published a "Theory of the Skull" embodying such a conception; and since [540] the excellent Dumeril allowed a like hypothesis to be strangled in the birth by the small wit of a French academician.
The progress of modern science is so rapid, that one is unaccustomed to see half a century elapse after the promulgation of a doctrine, which is capable of being tested by readily accessible facts, without either its firm establishment or its decisive overthrow. But nevertheless, at the present day, the very questions regarding the composition of the skull, which were mooted and discussed so long ago by the ablest anatomists of the time, are still unsettled; the theory of the vertebrate skull is one of the most difficult and, apparently inextricably confused subjects, which the philosophic anatomist can attack, and in consequence, not a few workers in science look, somewhat contemptuously, upon what they are pleased to term mere hypothetical views and speculations.
Indeed, though the germ of a great truth did really lie in these same hypotheses, its late or early development into a sound, and consequently fruitful, body of doctrine depended upon the manner in which biologists set about solving the problem presented to them; upon the clearness with which they apprehended the nature of the questions they wished to put, and the consequent greater or less fitness of the method by which their interrogation of nature was conducted.
I apprehend that it has been and is, too often forgotten that the phrase "Theory of the Skull" is ordinarily employed to denote the answers to two very different questions: the first, Are vertebrate skulls constructed upon one and the same plan?–second, Is such plan, supposing it to exist, identical with that of the vertebral column?
It is also forgotten that, to a certain extent, these are independent questions; for though an affirmative answer to the latter implies the like reply to the former, the converse proposition by no means holds good; an affirmative response to the first question being perfectly consistent with a negative to the second.1
As there are two problems, so there are two methods of obtaining their solution. Employing the one, the observer compares together [541] a long series of the skulls and vertebral columns of adult Vertebrata, determining, in this way, the corresponding parts of those which are most widely dissimilar, by the interpolation of transitional gradations of structure. Using the other method, the investigator traces back skull and vertebral column in their earliest embryonic states, and determines the identify of parts by their developmental relations.
It were unwise to exalt either of these methods at the expense of its fellow, or to be other than thankful that more roads than one leads us to the attainment of truth. Each, it must be borne in mind, has its especial value and its particular applicability, though at the same time it should not be forgotten that to one, and to one only, can the ultimate appeal be made, in the discussion of morphological questions. For seeing that living organisms not only are, but become, and that all their parts pass through a series of states before they reach their adult condition, it necessarily follows that it is impossible to say, that two parts are homologous or have the same morphological relations to the rest of the organism, unless we know, not only that there is no essential difference in these relations in the adult condition, but that there is no essential difference in the course by which they arrive at that condition. The study of the gradations of structure presented by a series of living beings may have the utmost value in suggesting homologies, but the study of development alone can finally demonstrate them.
Before the year 1837, the philosophers who were occupied with the Theory of the Skull confined themselves, almost wholly, to the first-mentioned mode of investigation, which may be termed the "method of gradations." If they made use of the second method at all, they went no further than the tracing of the process of ossification, which is but a small, and by no means the most important part of the whole series of developmental phenomena, presented by either the skull or the vertebral column.
But between the years 1836 and 1839, the appearance of three or four remarkable Essays, by Reichert, Hallmann, and Rathke,2 inaugurated a new epoch in the history of the Theory of the Skull. Hallmann's work on the Temporal Bone is especially remarkable for the mass of facts which it contains, and for that clearness of insight [542] into the architecture of the skull, which enabled him to determine the homologies of some of the most important bones of its upper arch throughout the vertebral series. Rathke showed the singular nature of the primordial cranial axis, and Reichert pointed out in what way alone the character of its lower arches could be determined. For the first time, the student of the morphology of the skull was provided with a criterion of the truth or falsity of his speculations, and that criterion was shown to be Development.
My present object is to lay before you a brief statement of some of the most important results to which the following out of the lines of inquiry opened up by these eminent men seems to lead. Much of what I have to say is directed towards no other end than the revival and justification of their views–a purpose the more worthy and the more useful, since with one or two honourable exceptions–I allude more particularly to the recent admirable essays of Prof. Goodsir–later writers on the Theory of the Skull have given a retrograde impulse to inquiry, and have thrown obscurity and confusion upon that which twenty years ago had been made plain and clear.
I have said that the first question which offers itself is, whether all vertebrate skulls are or are not, constructed upon a common plan, and in entering upon this inquiry I shall assume (what will be readily granted), that if it can be proved that the same chief parts, arranged in the same way, are to be detected in the skulls of a Sheep, a Bird, a Turtle, and a Carp, the problem will be solved affirmatively, so far, at any rate, as the osseous cranium is concerned.
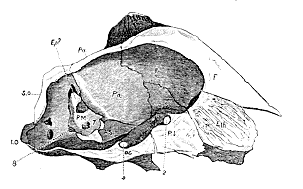
Composition of the Skull of a Sheep (fig. 1).
On examining a section of the cranium of a sheep, made either along a vertical and longitudinal, or a transverse and horizontal plane, a more or less completely ossified mass is observed in the middle line below, which forms part of the floor of the cranial cavity but extends beyond it. This may be termed the 'craniofacial axis.' Posteriorly it is a broad plate flattened from above downwards and is nearly parallel with the long axis of the cranial cavity; but from a point immediately behind the sella turcica, it becomes thicker and is compressed from side to side, so that, at the anterior boundary of the sella turcica, the craniofacial axis is much deeper than wide, and assumes the form of a vertical plate. From the anterior boundary of the cranial cavity onwards, or in its facial portion, the axial plate is very deep and very thin, and a line drawn through its longitudinal [543] axis would cut that of the cranial cavity at a very considerable angle. The craniofacial axis then is naturally divisible into three regions; a middle thick part, lodging the sella turcica, and composed of the basisphenoid behind and presphenoid in front, the two being separated by a suture; a posterior, lamellar, horizontally-flattened part, forming in the young animal a distinct bone, the basioccipital, bounding the occipital foramen behind and uniting with the basisphenoid in front; and an anterior laterally compressed portion, composed of the bony "lamina perpendicularis" of the ethmoid above and behind, united by the cartilaginous septum narium to the bony vomer below. This anterior division of the axis may be termed its ethmovomerine portion. Its posterior edge helps to close the anterior outlet of the cranial cavity, from which it is otherwise completely excluded.
Fig. 1.–Longitudinal section of the Skull of a Sheep. In this and the following sections of Crania the letters have the same meaning.
B.O. Basioccipital. A.S. Alisphenoid. Foramina for nerves. B.S. Basisphenoid. O.S. Orbitosphenoid. 1. Olfactory; 2. optic; 3. & P.S. Presphenoid. Pf. Prefrontal. 4. oculomotor and pathetic Eth. Ethmoid (lamina Sq. Squamosal. nerves; 5. third division of per-pendicularis). Ep. Epiotic. trigeminal; 7. portio dura and E.O. Exoccipital. S.O. Supraoccipital. mollis; 8. pneumogastric; M. Mastoid. Pa. Parietal. Epiph. Pineal gland, or P. or P.S. Petrosal. F. Frontal. epiphysis cerebri. P.M. Petromastoid.
The sella turcica lodges the pituitary body, and the synchondrosial union between the basisphenoid and presphenoid is situated so far forwards that the anterior wall of the fossa is almost wholly formed by the rostrum-like anterior prolongation of the basisphenoid. The [544] spinal cord passes out behind the posterior margin of the basi-occipital. The olfactory nerves leave the skull on each side of the ethmovomerine division of the craniofacial axis.
The walls of the cranial cavity are formed by a number of bones, which are divisible into two series, a superior and a lateral. Of the latter, four pairs of bones, separated by natural lines of demarcation, or sutures, are distinguishable, three of which abut directly upon the cranio-facial axis, while the fourth pair are only indirectly connected with it. Behind are the exoccipitals,3 united with the basioccipital and forming the lateral boundaries of the occipital foramen. In front of these are the petromastoids, complex bones which contain the auditory labyrinth, and are connected with the anterior part of the basioccipital and the posterior and superior part of the basisphenoid, only by cartilage.
Next come the alisphenoids, which are attached to the inferoposterior and the anterior portions of the basisphenoid. And, lastly, the orbitosphenoids articulate with the upper margins of the vertically elongated presphenoid.
In the superior series only four bones can be counted, of which two are single and two are pairs. The hindermost is the supraoccipital bone. It articulates with both the exoccipitals and the petromastoids. The next, in front, is the parietal, single in the adult sheep, but composed of two symmetrical halves in the lamb. It articulates with the petromastoids and with the alisphenoids. The frontals, or anterior paired bones, lastly, unite with the orbitosphenoids, and, in front of them, with the ethmoid.
Most important relations exist between the contents of the cranium and these constituent elements of its walls. The par vagum makes its exit between the exoccipital and the petromastoid; the portio dura and portio mollis enter the petromastoid; the third division of the trigeminal passes through the large "foramen ovale," which, in the sheep, has the exceptional peculiarity of being situated nearly in the middle of the alisphenoid; the optic nerve passes through a foramen included between the orbito- and pre-sphenoids, while, as has been mentioned above, the olfactory nerve passes out beside the ethmoid and in front of the orbitosphenoid. The relation [545] of the pituitary body, or hypophysis cerebri, to the upper surface of the basisphenoid, has already been alluded to; it, of course, gives more or less nearly the position of the third ventricle and crura cerebri. A style passed horizontally through the gemina, or mesencephalon, would strike against, or close to, the anterior margin of the petromastoid bone.
On turning to the exterior of the skull, certain bones come into view which were before invisible, as they take no share in forming the lateral walls of the cranial cavity, but are as it were, stuck on to the outer surface of these walls. The principal of these is the great squamosal bone, applied to the outer surfaces of the petro-mastoid, parietal and alisphenoid bones, sending off its zygomatic process to unite with the jugal, and furnishing the articular surface for the condyle of the lower jaw.
Partly articulated with the squamosal and partly with the petro-mastoid, is the irregular capsule of the tympanic bone, to which the tympanic membrane is attached, on whose removal the ossicule auditûs come into view, consisting of the malleus, incus, and stapes. The processus gracilis of the first of these bones lies between the tympanic and the squamosal. The short process of the incus abuts against the inner wall of the tympanum, just below the squamosal and close to the line of junction of the petrous and mastoid. These are the leading points in the structure of the. sheep's cranium to which I wish to direct attention at present. Bearing them in mind, let us now proceed to the consideration of the skull of a bird.
Composition of the Skull of a Bird (fig 2).
In most adult birds, as is well known, the bones of the cranium have coalesced so completely as to be undistinguishable. But in the chick, and to a greater or less extent, in the adult struthious bird, the boundaries of the various bones are obvious enough; and I will therefore select for comparison with the mammalian skull that of an ostrich, and that of a young chicken.
The craniofacial axis of the bird has the same general figure as that of the sheep, consisting of a thick, solid, median portion, lodging the sella turcica; of a posterior, horizontally, and of an anterior, vertically, expanded division; but it is comparatively shorter and thicker in correspondence with the greater shortness, in proportion to its depth, of the cranial cavity. The sella turcica is very deep, and its front wall is very thick. The lower and anterior half of this wall is produced into a long tapering process, which extends forwards [546] far beyond the anterior limit of the bony lamina perpendicularis of the ethmoid, to end in a point.
Overlying this process, and articulated with more than the posterior half of its upper surface, there is, in the ostrich, a strong thick, vertical, bony plate, narrower in front and behind than in the middle, and below than above. A curved vertical ridge on each lateral surface marks the line of its greatest transverse diameter and seems to indicate a primitive division of the mass into two parts, an anterior and a posterior. The latter is connected above with the bony plates representing the orbitosphenoids. The former exhibits on each side, posteriorly and superiorly, a groove, in which the olfactory nerve rests and, above this, expands into an arched process, which supports the anterior extremity of the frontal bone. Anteriorly, the superior end of the bone widens into a rhomboidal plate, which appears externally between the nasal bones. These anterior and posterior processes of the superior edge of the bone are connected by a delicate ridge, which passes from one to the other above, but leaves an irregular oval gap below.
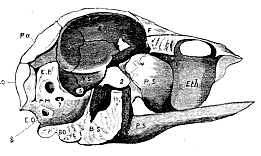
Fig. 2.–Longitudinal section of the Skull of a young Ostrich.
The anterior edge of the bony plate in question is continued into the unossified septum narium, which below supports the delicate bony representative of the vomer.
In the chick, the whole of the parts just described are unossified, but the composition and structure of the rest of the axis is essentially the same as in the ostrich.
It is not difficult to identify in the craniofacial axis of the birds parts corresponding with those which have been shown to exist in the mammal. In the chick, the basioccipital can be readily separated [547] from the basisphenoid. The latter has the same relation to the sella turcica in the bird as in the mammal; and only differs from it in that singular beaklike process, into which its inferior portion is prolonged anteriorly, and which is produced, according to Kölliker,4 by the coalescence with the basisphenoid of a distinct ossification, which is developed in the presphenoidal cartilage and partially represents the presphenoid of the mammal. The rest of the presphenoidal cartilage is more or less completely ossified, and appears to be represented in the ostrich by that part of the "vertical bony plate" which lies behind the curved ridge referred to above; while that part of the plate which is situated in front of the ridge, answers to the lamina perpendicularis of the ethmoid.
Nothing can be more variable, in fact, than the mode in which the ossification of the presphenoidal and ethmoidal portions of the craniofacial axis takes place in birds; while nothing is more constant than the general form preserved by these regions, and their relation to other parts, irrespectively of the manner in which ossification takes place in them. And in these respects birds do but typify the rest of the oviparous Vertebrate
If we compare the inferolateral walls of the ostrich's cranium with those of the sheep, we find the most singular correspondences. Posteriorly are the exoccipitals, which contribute to form the single condyloid head for articulation with the atlas, but otherwise present no important differences. In front of the exoccipital lies a considerable bony mass, which unites, internally and inferiorly, with the basioccipital and basisphenoid bones, and posteriorly is confluent with the exoccipitals. Its anterior margin is distinguishable into two portions, a superior and an inferior, which meet at an obtuse angle. The anterior inferior portion articulates with the alisphenoid; the anterior superior portion with the parietal. The anterior, posterior and inferior, relations of this bone are therefore the same as those of the petromastoid of the sheep.
Superiorly and posteriorly, a well-marked groove (which, however, is not a suture) appears to indicate the line of demarcation between the supraoccipital and this bone, whose pointed upper extremity appears consequently to be wedged in between the supraoccipital and the parietal.
The par vagum passes out between the bony mass under description and the exoccipital; the third division of the trigeminal leaves the skull between it and the alisphenoid. The portio dura and the portio mollis enter it by foramina very similarly disposed to those in [548] the sheep. Superiorly there is a fossa on the inner face of the bone which corresponds with a more shallow depression in the sheep, and, like it, supports a lobe of the cerebellum. Finally, the anterior inferior edge of the bone traverses the middle of the fossa which receives the mesencephalon. In every relation of importance, therefore, this bony mass corresponds exactly with the petromastoid of the sheep, while it differs from it only in its union with the exoccipitals and the supraoccipital posteriorly, and its contact with the craniofacial axis below.
If from the ostrich we turn to the young chick (fig. 3), the condition of this part of the walls of the skull will be found to be still more instructive. The general connexions of the corresponding bony mass, Pt. M. Ep., are as in the ostrich; but while it is even more evident that the groove appearing to separate its upper end from the supraoccipital is no longer a real suture (whatever it may have been), a most distinct and clear suture, of which no trace is visible in the ostrich's skull, traverses the bone at a much lower point, dividing it into an inferior larger piece, united with the exoccipital, and a superior portion, anchylosed with the supraoccipital. The latter contains the upper portions of the superior and external semicircular canals.
Moreover, on endeavouring to separate the inferior bone from the exoccipital, it readily parts along a plane which traverses the fenestra ovalis externally, and the anterior boundary of the foramen of exit of the par vagum internally. The posterior smaller portion remains firmly adherent to the exoccipital, while the other larger portion comes away as a distinct bone.
The latter answers exactly to the mammalian petrosal, while the small posterior segment corresponds with the mammalian mastoid. Like that of the mammal, it is eventually anchylosed with the petrosal; but unlike that of the mammal, it is also, and indeed at an earlier period, confluent with the exoccipital.5
Thus, to return to the ostrich's skull, the bony mass interposed between the exoccipital, supraoccipital and parietal bones, and the craniofacial axis, is in reality composed of three bones, an anterior, petrosal, a posterior, mastoid, and a third, which is distinct from the petrosal and mastoid in the chick, but is anchylosed with them in the ostrich, and which has as yet received no name. I shall term it, from its position with respect to the organ of hearing, the epiotic bone, "os epioticum."6
[549] The homology of the bone here called petrosal, with that of the mammal, is admitted by all anatomists. The bone which lies immediately in front of the petrosal is, with a no less fortunate unanimity, admitted to be the homologue of the mammalian alisphenoid. But is worthy of particular remark, in reference to the shifting of the relative positions of the lateral elements of the cranial wall, which has been imagined to take place in the ovipara, in consequence of the supposed invariable disappearance of the squamosal from the interior of their skulls; that although precisely the same bones are visible on the inner surface of the cranial cavity in the ostrich as in the sheep, the squamosal being absent in both, yet in the ostrich the third division of the trigeminal does not pass through the middle of the alisphenoid, but between it and the petrosal.7
The orbitosphenoids appear like mere processes of the presphenoid, and their relation to the optic nerves is altered in the same way (when compared with the corresponding bones in the sheep) as that of the alisphenoids to the trigeminal, that is to say the nerves pass behind, and not through them.
The superior series of bones in the cranial wall is exactly the same as in the sheep, and the parietals are distinct in the young ostrich, as in the lamb.
Attached to the exterior of the skull of the ostrich are, as in the sheep, several bones; but the appearance of some of these is widely different from that of the parts which correspond with them in the mammal. This is at least the case with the largest and uppermost of these bones, which lies upon the parietal above, the alisphenoid in front, and the exoccipital behind; while internally it is in relation with the petromastoid.
This bone lies immediately above an articular surface, which is furnished to the os quadratum by the petrosal, and more remotely it helps to roof in the tympanic cavity but takes no share in the formation of the fenestra ovalis. It sends a free pointed process downwards and forwards, which does not articulate with the jugal. Except in this particular, however, the bone in question resembles in every essential relation the squamosal of the sheep, while to the same extent it differs from the mastoid of that animal.
I have stated that in the ostrich this bone does not appear upon the inner surface of the wall of the skull, and in this respect, while it resembles the squamosal of the sheep and Ruminants generally, it differs from that of most other Mammalia, in which the squamosal makes its appearance in the interior of the skull, between the parietal, [550] frontal, alisphenoid and petrosal bones, and so contributes more or less largely to the completion of the cranial wall.
But it has been most strangely forgotten that the relations of the bone in question in birds, are by no means always those which obtain in the ostrich. In the young of the commonest and most accessible of domestic birds, in the chicken, the squamosal may be readily seen to enter largely into the cranial wall; a rhomboidal portion of its anterior and internal surface being interposed in front of the petrosal, between this bone, the parietal, the frontal, and the alisphenoid (Sq. fig. 3).
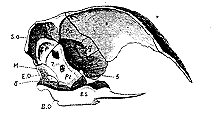
Fig. 3.–Longitudinal section of the Skull of a young Chicken.
There is therefore not a single relation (save the connexion of the jugal) in which this bone does not resemble the squamosal of the Mammalia–there is not one in which it does not differ from their mastoid.
The second bone applied externally to the cranium in the bird, is that large and important structure, the os quadratum, which intervenes between the petrosal and squamosal bones above, and the articular portion of the lower jaw below; which articulates with the pterygoid internally, and with the quadratojugal externally, which gives attachment to a part of the tympanic membrane, posteriorly, and which is very generally termed the tympanic bone, from its supposed homology with the bone so named in the Mammalia. The resemblance to the tympanic bone, however, hardly extends beyond its relation to the tympanic membrane; for in no other of the particulars mentioned above do the connexions of the two bones correspond. The tympanic of the mammal does not articulate with the lower jaw, nor with the pterygoid,8 nor with the jugal or quadratojugal. On the other hand, if the connexions of the tympanic membrane were sufficient to determine the point, not only the quad[551]ratum, but the articular element of the lower jaw, and even some cranial bones, must be regarded as tympanic.9
Again, if we trace the modifications which the tympanic bone undergoes in the mammalian series, we find that in those mammals, such as Echidna and Ornithorhynchus, which approach nearest to the Ovipara, and which should therefore furnish us with some hint of the modifications to which the tympanic bone is destined in that group, the bone, so far from increasing in size and importance, and taking on some of the connexions which it exhibits in the oviparous Vertebrata, absolutely diminishes and becomes rudimentary, so that the vast bony capsule of the placental mammal is reduced, in the monotreme, to a mere bony ring.
But it is no less worthy of remark, that in these very same animals the malleus and incus have attained dimensions out of all proportion to those which they exhibit in other mammals, and that they even contribute to the support of the tympanic membrane.
So far, therefore, from being prepared by the study of those Mammalia which most nearly approach the Ovipara, to find, in the most highly organised of the latter, an immense os tympanicum, with a vanishing malleus and incus, we are, on the contrary, led to anticipate the disappearance of the tympanicum, and the further enlargement of the ossicula auditûs. Thus far the cautious application of the method of gradations leads us, and leads us rightly–though the demonstration of the justice of its adumbrations can only be obtained by the application of the criterion of development.
It is twenty-one years since this criterion was applied by Reichert. Since his results were published, they have been, in their main features, verified and adopted by Rathke, the first embryologist of his age; and yet they are ignored, and the quadratum of the bird is assumed to be the tympanic of the mammal, in some of the most recent, if not the newest discussions of the subject. Reichert and Rathke have proved, that in the course of the development of either a mammal or a bird, a slender cartilaginous rod makes its appearance in the first visceral arch, and eventually unites with its fellow, at a point corresponding with the future symphysis of the lower jaw. Superiorly, this rod is connected with the outer surface of the cartilage, in which the petrosal bone subsequently makes its appearance. Near its proximal end, the rod-like "mandibular cartilage" sends off another slender cartilaginous process, which extends forwards parallel with the base of the skull. With the progress of development, ossification takes place in the last-named cartilage, and [552] converts it, anteriorly, into the palatine, and posteriorly, into the pterygoid bone. The mandibular cartilage itself becomes divided into two portions, a short, proximal, and a long, distal, by an articulation which makes its appearance just below the junction of the pterygopalatine cartilage. The long distal division is termed, from the name of its original discoverer, Meckel's cartilage. It lengthens, and an ossific deposit takes place around, but, at first, not in it. The proximal division in the mammal ossifies, but usually loses its connexion with the pterygoid, remains very small and becomes the incus.
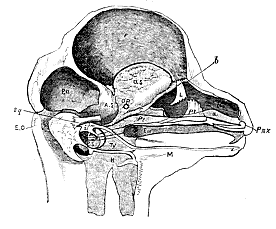
Fig. 4.–Dissection of the cranium and face of a Fœtal Lamb 2 in. long. The letters have the same signification as elsewhere, except N. Nasal capsules. a. b. c. Septum narium. L. Lacrymal. Pl. Palatine. Eu. Arrow indicating the course of the Eustachian tube. i. Incus. m. Malleus. M. Meckel's cartilage. H. Hyoid. Ps. Petrosal. Ty. Tympanic.
In the bird the corresponding part enlarges, ossifies, and becomes the os quadratum, retaining its primitive connexion with the pterygoid. In the mammal, the proximal end of Meckel's cartilage ossifies and becomes the malleus, while the rest ultimately disappears. The ossific mass which is formed around Meckel's cartilage remains quite distinct from the proximal end of that cartilage, or the malleus gradually acquires the form of the ramus of the lower jaw, and eventually developes a condyle which comes into contact and articulates with, the squamosal. In the bird, on the contrary, the ramus [553] of the jaw unites with the ossified proximal end of Meckel's cartilage which becomes anchylosed with the ramus, but retaining its moveable connexion with the quadratum (or representative of the incus), receives the name of the articular piece of the jaw. The rest of Meckel's cartilage disappears.
Thus the primitive composition of the mandibular cartilaginous arch is the same in the bird as in the mammal; in each, the arch becomes subdivided into an incudal and a Meckelian portion; in each, the incudal and the adjacent extremity of the Meckelian cartilage, ossify, while the rest of the cartilaginous arch disappears and is replaced by a bony ramus deposited round it. But from this point the mammal and the bird diverge. In the former, the incudal and Meckelian elements are so completely applied to the purposes of the organ of hearing, that they are no longer capable of supporting the ramus, which eventually comes into contact with the squamosal bone. In the latter, they only subserve audition so far as they help to support the tympanic membrane, their predominant function being the support of the jaw.
The tympanic bone of every mammal is, at first, a flat, thin, curved plate of osseous matter, which appears on the outer side of the proximal end of Meckel's cartilage, but is as completely independent of it as is the ramus of the jaw of the rest of that cartilage. In most birds it has no bony representative.10
It is clear, then, as Professor Goodsir11 has particularly stated, that the os quadratum of the bird is the homologue of the incus of the mammal, and has nothing to do with the tympanic bone; while the apparently missing malleus of the mammal is to be found in the os articulare of the lower jaw of the bird.
It would lead me too far were I to pursue the comparison of the bird's skull with that of the mammal further. But sufficient has been said, I trust, to prove that, so far as the cranium proper is concerned there is the most wonderful harmony in the structure of the two, not a part existing in the one which is not readily discoverable in the same position, and performing the same essential functions, in the other. I have the more willingly occupied a considerable time in the demonstration of this great fact, because it must be universally admitted that the bones which I have termed petrous, squamosal, mastoid, quadratum, articulare in the bird, are the homologues of particular bones in other oviparous Vertebrata, and consequently, if [554] these determinations are correct in the bird, their extension to the other Ovipara is a logical necessity. But the determination of these bones throughout the vertebrate series is the keystone of every theory of the skull–it is the point upon which all further reasoning must turn; and therefore it is to them, in considering the skulls of the other Ovipara, that I shall more particularly confine myself.
Composition of the Skull of the Turtle
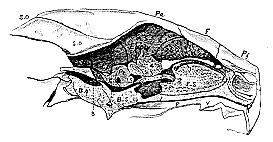
Fig. 5.–Longitudinal section of the Skull of a Turtle (Chelone mydas), exhibiting the relations of the brain to the cranial walls. The dotted parts marked AS. OS. PS. and Eth. are cartilaginous.
It has been seen that in birds the presphenoid, ethmoid, and orbitosphenoid regions are subject to singular irregularities in the mode and extent of their ossification. In the turtle, not only are the parts of the cranium which correspond with these bones unossified, but its walls remain cartilaginous for a still greater extent. In fact, if a vertical section be made through the longitudinal axis of a turtle's skull, it will be observed that a comparatively small extent of the cranial wall, visible from within, is formed by bone, and that the large anterior moiety is entirely cartilaginous and unossified. The anterior part of the posterior, bony, moiety of the cranial wall is formed by a bone (Pt.), whose long, vertical, anterior-inferior margin forms the posterior boundary of the foramen by which the third division of the trigeminal nerve makes its exit from the skull. The anterior and superior margin of the bone is very short, and articulates with the parietal bone. The superior margin is inclined backwards and articulates with the supraoccipital The posterior margin is straight, and abuts against a cartilaginous plate interposed between [555] this bone and that which succeeds it. The inner face of the bone is, as it were, cut short and replaced by this cartilage, whence the inferior edge is also short and is connected only with the basisphenoid, and not with the basioccipital. The anterior margin of the bone corresponds with the middle of the mesencephalon, while its inner face presents apertures for the portio dura and portio mollis. The posterior margin of its outer face forms half the circumference of the fenestra ovalis, and it contains the anterior and inferior portions of the labyrinth. Thus, with the exception of the absence of an inferior connexion with the basioccipital,–a circumstance fully explained by the persistence in a cartilaginous state of part of the bone, it corresponds in the closest manner with the petrosal of the bird. I confess I cannot comprehend how those who admit the homology of the bone called petrosal in the bird with that called petrosal in the mammal (as all anatomists do), can deny that the bone in question is also the petrosal, and affirm it to be an alisphenoid. The general adoption of such a view would, I do not hesitate to say, throw the Theory of the Skull into a state of hopeless confusion, and render a consistent terminology impossible. Where then is the alisphenoid? I reply, that it is unossified. The posterior portion of the cartilaginous side-wall of the skull, in fact, unites with the parietal, the petrosal, and the basisphenoid, just in the same way as the bony alisphenoid of the bird unites with those bones. Furthermore, as in the bird, it bounds the foramen for the third division of the trigeminal nerve anteriorly, and is specially perforated by the second division of the fifth, while the optic and the other divisions of the fifth pass out in front of or through its anterior margin.
Not only is the alisphenoid cartilaginous, but the orbitosphenoid is in the same condition, and a great vertical plate of cartilage represents the whole anterior part of the craniofacial axis, or the presphenoid and ethmovomerine bones.12 It has been imagined, indeed, that the rostrum-like termination of the basisphenoid represents the presphenoid, but I think this comes of studying dry skulls. Those who compare a section of the fresh skull of a turtle with the like section of the skull of a lamb, will hardly fail to admit that the rostrum of the basisphenoid in the turtle is exactly represented by that part of the sheep's basisphenoid, which forms the anterior and inferior boundary of the sella turcica, and that the suture between the basisphenoid and the presphenoid in the sheep corresponds [556] precisely with the line of junction between the rostrum of the basisphenoid and the presphenoidal cartilage in the turtle.
Connected with the posterior edge of the petrosal by the cartilaginous plate, which has been referred to above, and between this and the exoccipital, there appears, on the inner aspect of the longitudinal section of the turtle's skull, a narrow plate of bone connected above, with the supraoccipital, behind, with the exoccipital, below, with the basioccipital, and leaving between its posterior margin and the exoccipital an aperture whereby the par vagum leaves the skull. In fact, except in being separated from the petrosal by cartilage, this bone presents all the characters of the mastoid of the bird, which it further resembles in forming one-half of the circumference of the fenestra ovalis. In other respects it is more like the mastoid of the sheep, for it is not anchylosed with the exoccipital; it is produced externally into a great bony apophysis, which gives attachment to the representative of the digastric muscle; and it is largely visible external to the exoccipital, when the skull is viewed from behind. Indeed, the resemblance to the mastoid of the mammal is more striking than that to the corresponding bone in the bird. And I think it is hardly possible for any unprejudiced person to rise from the comparison of the chelonian skull with that of the mammal, with any doubt on his mind as to the homology of the two bones.
When the sheep's skull is viewed from behind, the posterior half of the squamosal is seen entering into its outer boundary above the mastoid. On regarding the turtle's skull in the same way, there is seen, occupying the same position, the bone which Cuvier, as I venture to think, most unfortunately, named "mastoid." But if the arguments brought forward above be, as I believe with Hallmann, they are, irrefragable, this bone cannot be the mastoid; and I can discover no valid reason why it should not be regarded as what its position and relations naturally suggest it to be–the squamosal. Its connections with the mastoid, petrosal, and quadratum are essentially the same as those of the squamosal in the bird and the mammal. The quadratum and articulare of the turtle are on all hands admitted to be the homologues of the similarly-named bones in the bird, and therefore all the reasonings which applied to the one apply to the other. When the petrosal, mastoid, and squamosa are determined in the turtle, they are determined in all the Reptilia. But the Crocodilia, Lacertilia, and Ophidia differ from the turtle and Chelonia generally, in that their mastoid is, as in the bird, anchylosed with the exoccipital. The squamosal, again, which in the Crocodilia essentially resembles that of the turtle, becomes a slender and elon[557]gated bone in the Lacertilia, and still more in the Ophidia, in which the quadratum is carried at its extremity.13
In the Amphibia the petrous and mastoid have the same relations as in the Reptilia; but it is interesting to remark, that in some Amphibia the anterior margins of the petrosal encroach upon the lateral walls of the skull so as completely to enclose the exit of the trigeminal, just as the posterior margin of the alisphenoid encroached so as to inclose it, in the sheep. It can be hardly necessary to remark, however, that this result has nothing to do with the disappearance of any element in the postero-lateral cranial walls, which have the same composition in the frog as in the crocodile or lizard.
The determination of the homologues of the squamosal, incudal, Meckelian, and tympanic elements in the amphibian skull is by no means an easy matter, but one requiring a much more careful investigation than it has yet received.
In Mammalia, a second arch, the hyoid, is connected with the outer surface of the skull, immediately behind the mandibular, and more particularly with that of the mastoid bone or its rudiment. The proximal end of this arch (which is, at first, like the mandibular arcade, a simple cartilaginous rod), in fact, usually becomes continuously ossified with the mastoid, forming part of the walls of the styloid canal; while below this, and external to the tympanum, it is converted into that slender bone, which is known as the styloid process.
In adult birds and most reptiles, the upper end of the hyoid arch is free, but in some Reptilia 14 it is attached by a styloid process to the representative of the mastoid. Whether attached to the cranium or not, in all abranchiate Vertebrata the proximal end of the hyoidean arch is quite distinct from that of the mandibular arch.
In the Amphibia, however, I find a condition of the proximal ends of these two arches, which seems to foreshadow that intimate connexion between them which obtains in fishes. On the outer side of the petrosal, and of that part of the exoccipital which represents the mastoid, there lies a cartilaginous mass, which is continued downwards into a pedicle, with whose lower end the mandible is articulated. From the anterior edge of the proximal half of this pedicle, the narrow cartilaginous basis of the pterygoid passes forwards and [558] upwards, to become directly continuous with the palatine bone in the frog, but to stop short of that point in the newt. Posteriorly, close to its proximal end, the pedicle becomes connected by a slender, fibrous or fibro-cartilaginous ligament with the upper extremity of the cornu of the hyoid. The hyoid and the mandibular arches are thus suspended to the skull by a common peduncle, which, to avoid all theoretical suggestion, I will simply term the "suspensorium."
The extent of the ossification which takes place, in and about this primitively cartilaginous suspensorium, varies greatly in different genera of Amphibia. Sometimes its distal end remains wholly unossified; sometimes, as in the common frog, a small outer portion of its lower extremity is ossified and sends a process forwards, becoming what is termed the quadratojugal bone; sometimes, as in the Triton, the distal half of the cartilage becomes more or less completely enclosed in a bony mass.
Another ossific deposit usually takes place in the outer half of the proximal end of the suspensorium, extending for a greater or less distance down towards the distal end, which it may even completely reach. It may be a simple triangular plate, as in Triton, or a T-shaped bone, as in Rana. In either case its lower end is the narrower, and fits into a kind of groove in the posterior and outer margin of the distal ossification.
This bone was considered by Cuvier to be the equivalent of the tympanic and the temporal (=squamosal); by Dugès it was called "temporomastoid."
The last constituent of this region of the skull in the Amphibia is one which is frequently overlooked altogether. In the frog, the membrana tympani is supported by a well-defined cartilaginous and partially ossified hoop, which is originally quite distinct from any of the elements of the suspensorium which have just been described, and which clearly deprives any of them of the right of being considered the homologue of the "tympanicum" of Mammalia.
I must defer the attempt to decide what the parts of the suspensorium really are, until the Piscine skull has been under consideration.
Composition of the Skull of the Carp.
The skulls of fishes present difficulties which necessitate, even for my present limited purpose, the entering into greater detail regarding them, than respecting those of the Reptilia or Amphibia. I select the cranium of the carp for description, as it departs far less widely [559] from the common plan, and therefore forms a better type for comparison with the skulls of other Vertebrata than that of any acanthopterygian or ganoid fish.
The craniofacial axis presents only four distinguishable bones. Behind, is the short basioccipital, with its cup for articulation with the first vertebra of the spinal column. In front of this is a greatly elongated bone, which, as in the bird, sends a process as far as the vomer, and forms the greater part of the axis of the skull; and which, I believe, represents, as in the bird, the basisphenoid and more or less of the presphenoid. The short vomer terminates the craniofacial axis anteriorly, and bears upon its upper surface a vertical septum, which, as in the bird, expands into a broad plate above, and is the ethmoid.
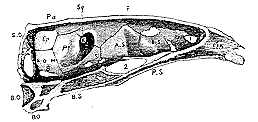
Fig. 6.–Longitudinal section of the Skull of a Carp (Cyprinus carpio).
The orbitosphenoids, united below, spring from the upper and anterior part of the presphenoid. Behind them the lateral walls of the skull are formed by the alisphenoid. These bones have the same essential relations as in the bird, for the olfactory nerves pass out of the skull over, and in front of, the orbito-sphenoids; the optic nerves make their exit behind and beneath these and the alisphenoid, while the trigeminal makes its exit behind the posterior edge of the alisphenoid. When viewed from within, the foramen ovale is seen to be as in the bird, a mere conjugational foramen between the alisphenoid and the bone which follows it; and on an external view, the third division of the trigeminal is seen to pass entirely in front of the last-named bone.
The minutest scrutiny of the relations of this bone only strengthens the conviction suggested by the first view of it, that it is the homologue of the petrosal of birds, and therefore of mammals and reptiles. As in the bird, the anterior margin of the fish's petrosal is divided into a superior and an inferior portion, which meets at an angle, [560] the superior portion articulating with the parietal (and squamosal), the inferior with the alisphenoid. Inferiorly, the petrous articulates with the basisphenoid, and, to a small extent, with the basioccipital. Posteriorly it articulates with a bone through which the pneumogastric passes, and which, guided by the analogy of most Reptilia, of Amphibia, and of birds, I believe to represent the coalesced or connate mastoid and exoccipital. The bone lodges the anterior part of the auditory labyrinth; its middle region corresponds with the middle of the mesencephalon. But as it does not separate the auditory organ from the cavity of the skull, it naturally presents no foramina corresponding with those through which the portio dura and portio mollis pass in abranchiate Vertebrata and Amphibia. There is one relation of the petrosal in the fish, however, in which it seems to differ from that of any of the oviparous Vertebrata hitherto described. Superiorly and posteriorly, in fact, it does not unite with the supraoccipital, which is small, comparatively insignificant, and occupies the middle of the posterior and superior region of the skull; but with a large and distinct bone which forms the internal of the two posterolateral angles of the skull, unites internally with the supraoccipital, anteriorly with the parietal and petrosal, inferiorly with the conjoined mastoid and exoccipital. It is the bone which was called "occipital externe" by Cuvier; and he and others have supposed it to be the homologue of that bone in the turtle which, following Hallmann, I have endeavoured to prove to be the mastoid. As I have already shown, the true mastoid of the fish must be sought elsewhere, and consequently the Cuvierian determination is inadmissible. And I must confess, that if our comparisons be confined to adult Vertebrata, the only conclusion which can be arrived at seems to be, that this bone is peculiar to fishes.
But a remarkable and interesting observation of Rathke, combined with the peculiar structure of the skull of the chick described above, leads me to believe that when their development is fully worked out, we shall find a distinct representative of this bone in many, if not all, vertebrate crania.
In his account of the development of Coluber natrix (see Note IV.), Rathke states that three centres of ossification make their appearance in that part of the cartilaginous wall of the cranium which immediately surrounds the auditory labyrinth. One of these is anterior, and becomes the petrosal; one is posterior, and eventually unites with the exoccipital; the third is superior, and in the end coalesces with the supraoccipital. The posterior ossification clearly [561] represents the mastoid, and it is most interesting to find it, in this early condition, as distinct as in the Chelonian.
The superior ossification has only to increase in size and remain distinct in the same way as the mastoid of the turtle remains distinct, to occupy the precise position of the "occipital externe" of the fish. But, further, it is most important to remark, that when this primarily distinct bone has coalesced with the supraoccipital, it stands in just the same relation to that bone, to the petrosal, to the mastoid and to the semicircular canals, in the snake, as that lateral element, early confluent or connate with the supraoccipital in the chick, which I have termed the "os epioticum." I believe, then, that this "os epioticum," distinct in the young snake, but afterwards confluent with the supraoccipital, and becoming what may be termed the epiotic ala of that bone in the adult, is the homologue of the corresponding bone, or confluent ala of the supraoccipital, in birds and reptiles, while in the fish it remains distinct, and constitutes the "occipital externe."
For the rest, the superior part of the cranial arch in the carp resembles that of the bird. There are a supraoccipital, two parietals, and two frontals; the squamosal occupies the same position as in the chick, and as in the latter, is, in the dry skull, visible from within, in front of the petrosal.
As in the Amphibia, both the mandibular and the hyoidean arches are suspended by a pedicle or suspensorium, which is, to a certain extent, common to both, and presents a complexity of structure which can only be elucidated by the most careful study of development.
In ordinary fishes, such as the carp, stickleback, &c., the proximal end of the suspensorium is constituted by a single bone, Cuvier's "temporal," whose cranial end abuts against the squamosal, petrosal, and post-frontal bones.
This temporal15 gives off posteriorly a process to which the cornu of the hyoid arch is attached; anteriorly and distally it ends in an expanded plate, with which two bones are connected, in front the tympanal, behind the symplectique. The distal end of the suspensor is constituted by the triangular jugal, whose distal and narrower extremity furnishes the condyle with which the mandible is articulated.
The elongated styliform symplectique is received into a groove on the posterior part of the inner surface of the jugal, and extends nearly to the condyle. In front, the jugal articulates with the transverse, [562] and more or less with the pterygoidien, which again are anteriorly connected with the palatine. The flat tympanal is fitted in between the pterygoidien, jugal, and temporal.
Besides these numerous bones, there are four others which enter less directly into the composition of the suspensorium. These are the pre-opercule, a sort of splint-like bone which lies on the outer and posterior faces of the temporal and jugal, and binds the two together; the operate, which articulates with a special condyle developed for it from the posterior edge of the temporal, above the attachment of the hyoid; the sousopercule, which lies in the opercular membrane beneath this; and lastly, the interopercule, the lowest of all, and commonly more or less closely connected with the angle of the lower jaw.
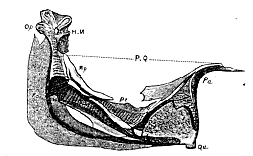
Fig. 7.–Palatosuspensorial arch of Gasterosteus from the inner side. HM. Hyomandibular bone. Op. Its articular facet for the operculum. Po. Pre-operculum. H. Articular surface for the styloid bone. Sy. Symplectic. P.Q. Palatoquadrate arch. Pa. Palatine bone. Qu. Quadratum. Pt. Pterygoid. Mp. Metapterygoid.
On examining the region in which these bones are eventually found, in an embryonic fish, I discovered, in their place, a delicate inverted cartilaginous arch, attached anteriorly, by a very slender pedicle, to the angles of the ethmoidal cartilage, and posteriorly connected by a much thicker crus with the anterior portion of that part of the cranial wall which encloses the auditory organ (fig. 8).
The crown of the inverted arch exhibits an articular condyle for the cartilaginous rudiment of the mandible. The posterior crus is not, as it appears at first, a single continuous mass, but is composed of two perfectly distinct pieces of cartilage applied together by their edges. The anterior of these juxtaposed pieces is continuous below [563] with the condyle-bearing crown of the arch, and with its anterior crus or pedicle (P.Q.). It is inclined backwards and upwards, and terminates close to the base of the skull in a free pointed extremity.
The posterior piece (S.Y.H.M.), on the other hand, has its broad and narrow ends turned in the opposite direction. Distally, or below, it is a slender cylindrical rod terminating in a rounded free extremity behind, but close to, the condyle for the mandible; above, it gradually widens and becomes connected with the cranial walls. On its posterior edge there is a convexity which articulates with the rudimentary operculum, and below this it gives off a short styloid process, to which the cartilaginous cornu of the hyoid is articulated. Thus the cartilaginous arch, which stretches from the auditory capsule to the ethmo-presphenoidal cartilage, consists, in reality, of two perfectly distinct and separate portions–the anterior division V-shaped, having its anterior crus fixed and its posterior crus free above; the posterior, styliform, parallel with the posterior leg of the V and free below. The anterior division supports the mandibular cartilage, the posterior the hyoidean cornu.
As ossification takes place, that part of the anterior crus of the V-shaped cartilage which is attached to the ethmo-presphenoidal cartilage becomes the palantine; its angle becomes the jugal ; between these two the transverse and pterygoidien (represented by only one bone in Gasterosteus) are developed in and around the anterior crus: the tympanal arises in the same way around the free end of the posterior crus. Thus these bones constitute an assemblage which is at first quite distinct from the other elements of the suspensorium, and immediately supports the mandibular cartilage.
The proximal end (H.M.) of the posterior styliform division gradually becomes articulated with the cranial walls, and, ossifying, is converted into the temporal . The distal cylindrical end (S.Y.) becomes surrounded by an osseous sheath, which at first leaves its distal end unenclosed. The bone thus formed is the symplectique, which is at first free, but eventually becomes enclosed within a sheath furnished to it by the jugal, and so strengthens the union of the two divisions of the arch already established by the junction of the tympanal with the temporal . The symplectique and temporal do not meet, but leave between them a cartilaginous space, whence the supporting pedicle of the hyoid, which ossifies and becomes the osselet styloide, arises.
The operculum, suboperculum, interoperculum, and preoperculum are not developed from the primitive cartilaginous arch, but make their appearance as osseous deposits in the branchiostegal membrane, behind, and on the outer side of, the posterior crus.
If we turn to the higher Vertebrata, we find, as I have stated above, that, at an early period of their embryonic existence, they also present a cartilaginous arch, stretching from the ethmo-presphenoidal cartilage to the auditory capsule, and supporting the mandibular or Meckelian cartilage on the condyle furnished by its inverted crown. The anterior part of the anterior crus of this arch becomes the palatine bone, which is therefore truly the homologue of the fishes' palatine. The posterior part of it becomes the pterygoid, which therefore is the homologue of the pterygoidien (and transverse ?) of the fish.
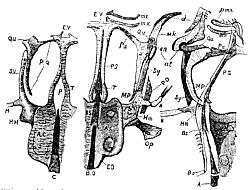
Fig. 8.–Cranium and face of young Gasterostei at different ages. The left-hand figure is a view of the base of the skull of a very young fish. The middle figure represents the under aspect, and the right-hand figure, a side view of a longitudinal section, of a more advanced stickleback's skull.
C. Notochord. P. Pituitary space. AC. Auditory capsules. T. Trabeculæ cranii. E.R. Ethmovomerine cartilage. P.Q. Palatoquadrate arch. Qu. Quadratum. S.Y. or So. Symplectic. H. Hyoidean arc. H.M. Hyomandibular cartilage. The other letters have the same signification as in the preceding figures, except pmx, Premaxilla. mx. Maxilla. d. Dentate. an. Angulare. at. Articulare. Mk. Meckel's cartilage.
The produced crown of the arch in the higher Vertebrata becomes either the incus, or its equivalent, the quadratum. I therefore entertain no doubt that the jugal is really the homologue of the quadratum of other oviparous Vertebrata. That the tympanal has no relation whatsoever with the bone of the same name in the [565] higher Vertebrata is indubitable; and I am unable to discover among them any representative of it. It seems to me to be an essentially piscine bone, to be regarded either as a dismemberment of the quadratum or of the pterygoid. It may be termed the "metapterygoid."
Still less do I find among the higher Vertebrata in their adult state, any representative of the posterior division of the suspensor, constituted by the temporal and symplectique. It is quite clear, that the temporal is not, as Cuvier's name would indicate, the homologue of the squamosal. The whole course of its development would negative such an idea, even if we had not a squamosal already; and I shall therefore henceforward term it, from its function of affording support to both the hyoid and mandibular arches, the hyomandibular bone, "os hyomandibulare," while the other bone of this division may well retain the name of symplectic.
It is commonly supposed that the hyomandibular, symplectic, metapterygoid, and quadrate are all to be regarded as mere subdivisions of the quadratum of higher Vertebrata. Such a view, however, completely ignores and fails to explain, the connexion of the hyoidean arch with the hyomandibular bone. In no one of the higher Vertebrata does such a connexion ever obtain between any part of the quadratum and the hyoid, which are quite distinct, and attached separately to the walls of the cranium, in even young embryos of the abranchiateVertebrata.
Nevertheless, in their very earliest conditions, these embryos are said to present a structure, which, if I mistake not, shadows forth the organization of the fish. The visceral arches, in which the mandibular and hyoid cartilages are developed, are at first separated to the very base of the cranium by a deep cleft, the anterior visceral cleft, so that the semi-cartilaginous rudiments of the mandibular and hyoid are completely separate. Subsequently they are said to coalesce above, as the visceral cleft diminishes, so as to have a common root of attachment to the cranium; and this, I apprehend, answers to the hyomandibular bone, and its prolongation to the symplectic. With advancing development, however, this part does not advance, but remains stationary, and becomes confounded with the wall of the cranium; so that the two arches subsequently appear to be attached to the latter quite independently, and there is nothing left to represent this division of the suspensorium in fishes.
I am strengthened in this view by the structure and development of the palatosuspensorial apparatus in the Amphibia, whose consideration I deferred when speaking of the skull in that class. [566] On examining a young tadpole (fig. 9), a cartilaginous process is seen to arise from the walls of the cranium, opposite the anterior part of the auditory capsule, and, passing obliquely downwards and forwards, to end in a rounded condyloid head, which articulates with the representative of Meckel's cartilage. At the anterior boundary of the orbit the process gives off a broad, nearly vertical apophysis (O), Which ends superiorly in a free, rounded, and incurved edge.
Fig. 9.–The upper right-hand figure represents a longitudinal section of the dissection of the head of a tadpole with external gills. The two lower figures represent dissections of the crania of tadpoles with well-developed hinder limbs. In the one, the integuments, organs of sense, &c. of the right side are taken away so as to lay bare the facial cartilages and the brain. In the other the cranium is opened from above, and the brain and myelon are extracted. The letters have the same signification as before, except My. Myelon. M. Mouth. olf. Olfactory sac. op. Eye. 1. Anterior cerebral vesicle. 2. Middle cerebral vesicle. 3. Posterior cerebral vesicle. Ia. Rhinencephalon. Ib. Prosencephalon. Ic. Deutencephalon, or vesicle of the third ventricle. I. II. III. IV. V. Branchial arches. x. Organs of adhesion. 1. Lips. 5. Trigeminal ganglion. 7. Ganglion of the portio dura. 8. Aperture for the exit of the pneumogastric.
The crotaphite muscle passes to its insertion on the inner side of this, the so-called "orbitar process." From the condyle the cartilaginous process sweeps upwards and inwards, and ends by passing into the ethmo-presphenoidal cartilage. It consequently forms an inverted arch, whose keystone is the condyle for Meckel's cartilage, and is, in its connexions and form, strictly comparable with the cartilaginous arch which I have described in the embryo fish. The posterior crus [567] of the arch, it is true, is not divided into two parts, but nevertheless it represents the whole suspensorium of the fish, and not merely the quadratum of the abranchiate vertebrate, because immediately behind the orbitar process it presents an excavated surface, which articulates with the proximal end of the cornu of the hyoid. That part of the cartilaginous arch, therefore, which lies above and behind this point, corresponds with the proximal division of the suspensorium in the fish, or with the hyomandibular bone; while that portion which lies below and in front of it, corresponds with the distal division of the suspensorium and the anterior crus of the arch in the fish, or in other words, with the symplectic, quadratum, metapterygoid, pterygoid, transverse, and palatine bones.
In the course of development, in fact, the palatine bone appears, as in the fish, in that part of the arch which is immediately connected with the ethmo-presphenoidal cartilage, and a single pterygoid in that part of its anterior crus which lies between the palatine and the articular portion, which obviously represents the quadratum. But this pterygoid is, in the adult frog, a large bone, which, on the one hand, stretches down on the inner side of the quadrate cartilage, and, on the other, sends a process inwards and upwards, which nearly reaches the base of the skull. If the pterygoid, transverse, and metapterygoid of the fish were anchylosed into one bone, or if the corresponding region of the primitive cartilage were continuously ossified, the result would be a bone perfectly similar to the pterygoid of the frog; and I entertain no doubt that the amphibian pterygoid does really represent these bones.
The inferior ossification in the batrachian suspensorium certainly answers to the quadratum, in Triton–whether it should be regarded partly or wholly as a quadrato-jugale in the frog seems to be a question of no great moment–inasmuch as we may be quite sure that the lower end of the frog's suspensorium represents the quadrate or incudal element in other vertebrate.
It is well known that, in the course of the development of the frog, the end of the suspensorium, as it were, travels backwards, so that its axis, instead of forming an acute angle, open forwards, with that of the cranium, as in the tadpole (fig. 9), forms a very obtuse angle, open downwards, in the adult frog. This change is accompanied by a relative and absolute lengthening of that part of the suspensorium which lies between the articulation of the hyoid and that of Meckel's cartilage (containing its proper quadrate portion), and by a relative shortening of that part which lies between the articulation of the hyoid and the skull (or the hyomandibular portion). The consequence of this is, that the articular surface for the hyoid appears constantly to approach the cranial wall, until at length, in the adults it seems to be almost in contact with it. If a knife were passed obliquely between the pterygoid and the suspensorium, and then carried through the suspensorium to its posterior margin a little above the condyle for the mandible, it would divide the suspensor into a proximal and a distal portion, precisely resembling those which naturally exist in the embryonic fish. If the proximal division ossified, it would clearly represent the hyomandibular and symplectic bones. Now in the Amphibia, although the suspensor is not thus divided, it ossifies very nearly as if it were, and the superior or proximal ossification is the so-called "temporo-tympanic," "temporo-mastoid," or "squamosal" bone.16
That this bone is really the homologue of the hyomandibular and symplectic in the fish, becomes, I think, still more clear when we compare it with such an aberrant form of piscine suspensorium as is presented by some of the eel-tribe (Muræna, e.g.). In these fishes the suspensorium is formed by only two bones, a small distal quadratum, which, as usual, articulates with the lower jaw, and a large wide proximal bone, which articulates above with the post-frontal and squamosal, gives attachment to the operculum and to the cornu of the hyoid, and sends down a process towards the articular head of the quadratum. The single bone, which represents the three pterygoids of other fishes, is articulated for the most part with the quadratum, but partly with this proximal bone. The latter, therefore, clearly represents both the hyomandibular and the symplectic bones of ordinary fishes.
But if the suspensorium of Triton be compared with that of Muræna, e.g., it will, I think, be hardly doubted, that while the distal ossification in the former corresponds with the quadrature the proximal answers (at any rate, chiefly) to the hyomandibular bone of the Muræna. Indeed it differs from the latter principally in being an ossific deposit in the outer portion only of the primitive cartilages.17
Thus it would seem, that in the manner in which the lower jaw is [569] connected with the cranium, Pisces and Amphibia, as in so many other particulars, agree with one another, and differ from Reptilia and Aves on the one hand, as much as they do from Mammalia on the other. And the difference consists mainly, as might be anticipated, in the large development in the branchiate Vertebrata of a structure which aborts in the abranchiate classes. A most interesting series of modifications, all tending to approximate the ramus of the mandible more closely to the skull,18 is observable as we pass from the fish to the mammal. In the first, the two are separated by the hyomandibular, the quadrate, and the articular elements, the first of which becomes shortened in the Amphibia. In the oviparous abranchiate Vertebrata the cranium and the ramus are separated only by the quadratum and the articulare, the hyomandibulare having disappeared. Finally, in the mammal, the quadratum and the articulare are applied to new functions, and the ramus comes into direct contact with the cranium.
The operculum, suboperculum, and interoperculum appear to me to be specially piscine structures, having no unquestionable representatives in the higherVertebrata. Much might be said in favour of the identification of the preoperculum with the tympanic bone; but there are many arguments on the other side, and at present I do not see my way to the formation of a definite conclusion on this subject.
In the preceding discussion of the structure of the osseous. vertebrate skull, I have desired to direct your attention, more particularly, to the consideration of those fundamental bones, the determination of whose homologues throughout the vertebrate series is of the greatest importance for my present object. The presphenoid, ethmoid, mastoid, and petrosal are the Malakhoff and the Redan of the theory of the skull; and if anatomists were once agreed about their homologues, there would be comparatively little left to dispute about.
But besides the axial, inferolateral, and superior series of bones there are other, less constant, elements of the cranial wall, forming a discontinuous superolateral series. These are the epiotic, the squamosal, the postfrontal, the prefrontal, and lacrymal bones. Of the two first-named of these bones I have already spoken sufficiently. The postfrontal exists only in Reptiles and Fishes, and is always situated between the frontal, alisphenoid, petrosal, and squamosal–[570] the extent to which it is absolutely in contact with any one of these bones varying.
The prefrontal and lacrymal bones are always developed in or upon that lateral process of the ethmosphenoidal plate, which gives attachment externally to the palatopterygoid arch; consequently they lie at the anterolateral ends of the frontal, and have more or less close relations with it, the ethmoid and the palatine bones.
Finally, the nasal bones (or bone) never enter into the composition of the walls of the skull, but have the same relation to the anterior and upper expanded edge of the prolonged lamina perpendicularis or body of the ethmoid, as the vomer or vomers have to its lower edge.
If the conclusions which I have laid before you are correct, the following propositions are true of all the bony skulls of Vertebrata.
1. Their axis contains at most five distinct bones, which are, from before backwards, the basioccipital, the basisphenoid, the presphenoid, the ethmoid, and the vomer; but any of these bones; except the basisphenoid, may be represented by cartilage, and they may anchylose to an indefinite extent; so that the number distinguishable as separate bones in any skull cannot be predicated. The craniofacial axis invariably presents the same regions, but the histological character of these regions may vary.
2. Their roof contains at most, leaving Wormian bones out of consideration, five bones (supraoccipital, parietals and frontals), or seven, if we include the epiotic bones in the roof. The number falls below this in particular cases, for the same reason as that given for the apparent variations in composition of the axis.
3. Their inferolateral wall contains at most six pair of bones (exoccipitals, mastoids, petrosals, alisphenoids, orbitosphenoids, prefrontals), whose apparent number, however, is affected by the same causes.
4. The axial bones have definite relations to the brain and nerves. The basioccipital lies behind the pituitary body, the basisphenoid beneath it, the presphenoid in front of it. In fact the pituitary body may be regarded as marking the organic centre, as it were, of the skull–its relations to the axial cranial bones being the same, as far as I am aware, in all Vertebrata.
The olfactory nerves pass on either side of the ethmoid, which bounds the cranial cavity in front, the greater part of its substance and that of the vomer being outside the cranial cavity.
5. The lateral bones have definite relations to the brain, nerves, and organs of sense. The exoccipital lies behind the exit of the par [571] vagum; the mastoid lies in front of it; the petrosal lies behind the exit of the third division of the trigeminal; the alisphenoid lies in front of it; though either bone may, to a certain slight extent, encroach on the province of the other. The optic nerve passes out more or less in front of the alisphenoid, and behind, or through, the orbitosphenoid.
The organ of hearing is always bounded in front by the petrosal bone, which limits the anterior moiety of the fenestra ovalis.
The organs of smell always lie on each side of the ethmovomerine part of the axis.
The greater part, or the whole, of the petrosal lies behind the centre of the mesencephalon.
6. The attachment of the mandibular arch to the skull is never situated further forward than the posterior boundary of the exit of the trigeminal; consequently it cannot belong to any segment of the skull in front of the petrosal.
But if propositions of this generality can be enunciated with regard to all bony vertebrate skulls, it is needless to seek for further evidence of their unity of plan. These propositions are the expression of that plan, and might, if one so pleased, be thrown into a diagrammatic form. There is no harm in calling such a convenient diagram the 'Archetype' of the skull, but I prefer to avoid a word whose connotation is so fundamentally opposed to the spirit of modern science.
Admitting, however, that a general unity of plan pervades the organization of the ossified skull, the important fact remains, that many vertebrated animals–all those fishes, in fact, which are known as Elasmobranchii, Marsipobranchii, Pharyngobranchii, and Ditnoi–have no bony skull at all, at least in the sense in which the words have hitherto been used. In these Vertebrata the skull is either membranous or cartilaginous; or if ossified, the ossific matter presents no regular grouping around a few distinct centres.
Thus the cranium of the Amphioxus is nothing but a membranous capsule, whose walls are continuous with those of the canal for the spinal cord, and in whose floor lies a continuation of the notochord which underlies the spinal canal.
In the Marsipobranchii there is a marked increase in the capacity of the cranium as compared with that of the spinal canal, in correspondence with the decided differentiation of the cerebral masses; and, at the same time, the cranial walls have undergone a more or less extensive chondrification. The notochord terminates in the midst of the firm and solid cartilaginous plate which forms the posterior part of the basis cranii, and which sends forward two processes, including a membranous interspace. The auditory capsules are enclosed within prolongations of the sides of the basilar plate; and just in front of and below them, the root of each process of the basal plate gives off a solid prolongation, which passes at first outwards and downwards, and then bends upwards and forwards, to rejoin the anterior part of the process of the basilar plate of its side. An inverted arch is thus formed, and the space included between its crura and the sides of the cranium, constitutes the floor of the orbit.
The posterior crus of the arch is divided into two, more or less distinct, pillars, the posterior of which supports the hyoidean arc; the mandibular arc appears to be absent.
The apertures whereby the cranial nerves make their exit are situated in the side-walls of the capsule, that for the vagus lying immediately behind the auditory capsule, while that for the trigeminal is immediately in front of the same organ. The olfactory nerves perforate the anterior walls of the cranial capsule; the optic, its lateral walls between them and the trigeminal.
The skulls of the Elasmobranchii, again, appear at first to be something quite different from either of these. The cranium is here a cartilaginous box, more or less incomplete and membranous above, and presenting on each side posteriorly a transverse enlargement, in which the auditory organ is contained; while anteriorly it expands into a broad plate, which on each side overhangs the olfactory sacs. The notochord and the membranous space have disappeared, or their traces only are visible in the base of the cranium, whose walls are, as it were, crusted with a multitude of minute plates of bone.
In the Chimæræ the inferolateral walls of the cranium pass into a cartilaginous arch-like plate which form the floor of the orbit, and: whose posterior part, as in the Marsipobranchii, gives attachment to the hyoidean arch; besides which, a mandibular cartilage is connected with the condyloid surface developed from the crown of the arched plate.
In the Plagiostomes there is also an inverted suborbitar arch with a mandibular cartilage and a hyoidean apparatus, but the structure of the arch is different from what obtains in Chimera.
The outer wall of that portion of the cranium which lodges the auditory organ, in fact, furnishes an articular surface for a strong moveable peduncle, to which the hyoid arc is usually attached. At its lower end, however, this peduncle does not articulate with the mandibular cartilage, but is directly connected with a strong cartilaginous plate which forms the upper boundary of the gape, and is articulated anteriorly with the sides of the skull in front of the orbit. [573] This plate bears the upper series of teeth, and bites more or less directly against the mandible, which is moveably articulated with a condyle furnished by its posterior extremity.
The upper plate is commonly, though, as I think, erroneously, regarded as the homologue of the maxilla and premaxilla in other fishes; the peduncle as the homologue of their whole suspensorium.19
The par vagum leaves the skull behind the auditory organ; the trigeminal passes out in front of it; and then its third division traverses the space enclosed between the peduncle, the upper plate, and the skull. The optic nerve passes through the lateral walls of the skull in front of the trigeminal, and the olfactory perforates its anterior boundary.
So brief and simple a statement of the characters of the skulls of these three orders of fishes, while it brings their diversities into prominence, also exhibits an amount of uniformity among them which is not a little remarkable. The exits of the great nerves have fixed relations to the auditory capsules, to the anterior boundary of the skull, and to the pituitary body. The inferior arc of the hyoid is constant (except in the Pharyngobrachii), and has always, speaking broadly, the same relative position with respect to the auditory capsule and the posterior crus of the suborbitar arch. The suborbitar arch itself is always present (except in Pharyngobranchii); its posterior crus is always attached to the cranium behind the third division of the trigeminal nerve, while the anterior is invariably fixed to that part of the skull which lies behind, or beside, the base of the olfactory capsule.
Thus the employment of the method of gradation alone exhibits a surprising uniformity in the organization of these lower forms of skull; and on comparing them with the higher forms, it seems obvious that, so far as it goes, their plan is identical with that of the latter; for the relations of the auditory organ to the par vagum and trigeminal are the same in each; the posterior crus of the suborbitar arch answers to the suspensorium of Teleostei, its anterior crus to their palatopterygoid apparatus. But with all this, there are discrepancies in the structure of the skull itself, which would forbid too close an approximation between the bony and the unossified crania, if their adult forms alone were examined. The study of the development of the ossified vertebrate skull, however, eliminates this difficulty, and satisfactorily proves that the adult crania of the lower Vertebrata are but special developments of conditions through which the embryonic crania of the highest members of the subkingdom pass.
[574] It is to Rathke's luminous researches that we are indebted for the first, and indeed, even now, almost the only, demonstrative evidence of this great fact. Twenty years ago that great and laborious embryologist worked out the early stages of the development of the skull in each class of the Vertebrata. Confirmed and adopted by Vogt and Bischoff, his conclusions have been feebly controverted, but never confuted; and my own observations lead me to believe that they are destined to take a permanent place among the data of biological science. Nothing is easier than to verify Rathke's views in an embryonic fish or amphibian; and as it matters not which of the higher Vertebrata is selected for the study of cranial developments I will state at some length what I have observed in the embryonic frog.20
Before the dorsal laminae have united so as to enclose the primitive craniospinal cavity, the anterior portion of the floor of that cavity is bent downwards. The angle which the deflexed portion forms with the rest becomes less and less obtuse, until, when the dorsal lamina have united and the visceral clefts have begun to appear, it constitutes a right angle.
On examining the floor of the craniospinal cavity at this period, it is seen that the notochord, at present formed by the aggregation of a number of yelk segments or embryo-cells, small in themselves, but larger than those of which the rest of the body is composed, ends in a point immediately behind the angular flexure.
The notochord has no sheath as yet, and is not in any sense prolonged into the deflexed portion of the floor of the craniospinal cavity.
When the visceral clefts first appear, they are best seen from the inner or pharyngeal aspect of the visceral wall. Five, of which the two anterior are the longest and about equal, while the others gradually diminish in length from before backwards, can be distinctly observed. They mark out the boundaries of a corresponding number of "visceral arches," and there is sometimes an appearance as of a sixth visceral arch behind the last cleft. A horizontal section shows that these arches differ in nothing but their relative size–in no other respect can one of them be distinguished from the other.
The anterior visceral cleft lies in a transverse plane, immediately behind the angular bend of the floor of the craniospinal cavity, or, as I shall henceforward term it, mesocephalic flexure. Consequently the posterior part of the first visceral arch passes into the future basis cranii close to the flexure.
The parts of the cerebrum are now distinguishable. It is bent in [575] correspondence with the mesocephalic flexure, and its most projecting portion, or the angle of the bend, is the rudiment of the mesencephalon. The large rudiment of the pituitary body lies immediately in front of the flexure, and is therefore altogether anterior to the end of the notochord and to the posterior part of the first visceral arch. The rudiment of the eye lies at first altogether in front of the flexure, and therefore anterior to the root of the first visceral arch.
The auditory vesicles make their appearance on each side of a line which would cut the chords a little behind its anterior termination. They are at first quite free and perfectly distinct from the walls of the cranium, which is in accordance with Remak's statement, that they are originally formed by the involution of the epidermic layer of the embryo. They long remain separate and easily detachable from the cranial walls.
Ten days after impregnation, larvæ with rudimentary external gills. and colourless blood, still exhibited some traces of the mesocephalic flexure, but the angle formed by the anterior and posterior portions of the cranium was very obtuse; the base of the cranium had, in fact, undergone a gradual straightening. The rudiments of the cranial skeleton had made their appearance, and consisted, behind the mesocephalic angle, of a broad semicartilaginous plate enclosing the anterior end of the notochord, but not covering it above or below. It is not as yet adherent to the auditory sacs.
That part of the middle of the basis cranii which underlies the pituitary body is not converted into cartilage, but remains membranous, and may be called the "subpituitary membrane." The delicacy of this membrane is so great that it is easily torn, when the pituitary body seems, as Rathke originally supposed, to unite with the palatine mucous membrane. But that this is not really the case, is readily demonstrable in an embryo whose tissues have been sufficiently hardened with alcohol or nitric acid.
The cartilaginous basal plate gives off a prolongation on either side of the subpituitary membrane. This, the "cranial trabecula" (Schädelbalke of Rathke), passes forwards with a slight convexity outwards, and then turning inwards comes into contact with its fellow (from which, however, it is at first distinct), and spreads out into a broad, flat, elongated process, which I shall term the ethmovomerine cartilage.
Behind the eye and just in front of the auditory capsule (in the posterior part of the first visceral arch, therefore), a cartilaginous process lies, which is connected proximally with the root of the [576] trabecula close to the basal plate, while at its distal end it sends a prolongation upwards to unite with the posterior end of the ethmovomerine cartilage. It then forms an arch, between which and the basis cranii is an interspace corresponding with, and lodging, the under surface of the large eyeball. The rudiments of the hyoid, mandibular and maxillary apparatus in larvæ at this stage are somewhat indistinct; and indeed not only in this, but in other respects, more instruction is to be derived from tadpoles which have advanced further.
In larvæ, with completely internal branchiae and very short tubercles in the place of hind limbs, the notochord suddenly narrows between the auditory capsules to hardly more than half its preceding dimensions, and then gradually tapers off, to what appears to be a rounded end, a short distance from the anterior boundary of the basal plate. On very careful examination, however, a delicate process (which may by possibility be nothing but a cavity in the cartilage) can be traced from it very nearly to the margin of the basal plate. But there is no continuation whatsoever, either of the notochord itself or of its sheath, into the subpituitary membrane, which is now composed of delicate connective tissue, and from its extreme thinness and transparency would exhibit the least trace of such a prolongation. And I speak the more confidently on this point, because the delicate process of the notochord or cavity in the cartilage, to which I have referred, contains opaque unchanged vitelline granules, and is therefore particularly conspicuous. The basal cartilage is still divided by the notochord into two lateral moieties, which are only united by a short band of cartilage in front of the end of the notochord. It sends off from its outer side a cartilaginous process, which envelopes the auditory capsule externally, but leaves on its inner side a wide aperture for the entrance of the auditory nerve. The oval auditory capsules thus formed have their long axis directed outwards and forwards.
The trabeculæ are still better developed than before, but instead of remaining distinct anteriorly, they have become fused together into a single trapezoidal cartilage, which may be termed the ethmo-presphenoidal plate. This plate, as it were, divides anteriorly into two flat, elongated and somewhat divergent processes, which are concave downwards and end in truncate extremities. Fibrous tissue connects the ends of these ethmovomerine processes with a crescentic cartilaginous plate which supports the horny upper jaw of the tadpole.
The posterior crus of the palatosuspensorial, or suborbitar, arch is [577] not yet united with that portion of the cranial wall which encloses the auditory capsule; but for the rest the same description applies to it which has already been given of the palatosuspensorial arch and its appendages in more advanced tadpoles. In this state, the roof, and all the lateral walls of the cranium, but that part into which the auditory capsule enters, are membranous.
If the skull of the larval frog just described, be laid open and the exit of the nerves observed (fig. 9), it will be seen that the par vagum makes its way out by a foramen situated immediately behind the auditory capsule; that the third division of the trigeminal leaves the cranium in front of the auditory capsule, passing over the posterior crus of the palatosuspensorial arch; and that the optic traverses the membranous walls of the skull between this and the olfactory nerve, which perforates the anterolateral region to enter the olfactory capsules. The latter are situated wide apart, on each side and in front of, the broad ethmo-presphenoidal cartilage and the anterior crus of the palatosuspensorial arch, and are even a little overlapped by the edges of the ethmovomerine processes.
In the further course of development, the trabeculæ approximate and elongate, so as to obliterate the subpituitary membrane, and form with the enlarged basal cartilage, the ethmoid cartilage and the ethmovomerine cartilages, the continuous cartilaginous craniofacial axis. A histological metamorphosis into cartilage is undergone by the roof of the occipital region of the skull, but in front of this it remains membranous; so that in the adult frog (in which this cartilaginous framework persists), the skull, when deprived of its bony matter, presents an anterior fontanelle. The ethmovomerine cartilages diverge still more, and form the broad mass whose lateral cavities shelter the olfactory sacs in the adult frog. If, bearing in mind the changes which are undergone by the palatosuspensorial apparatus and which have already been described, we now compare the stages of development of the frog's skull with the persistent conditions of the skull in the Amphioxus, the Lamprey, and the Shark, we shall discover the model and type of the latter in the former. The skull of the Amphioxus presents a modification of that plan which is exhibited by the frog's skull, when its walls are still membranous and the notochord is not as yet embedded in cartilage. The skull of the lamprey is readily reducible to the same plan of structure as that which is exhibited by the tadpole, while its gills are still external and its blood colourless. And finally, the skull of the shark is at once intelligible when we have studied the cranium in further advanced larvae, or its cartilaginous basis in the adult frog.
[578] Thus, I conceive, the study of the mode in which the skulls of vertebrate animals are developed, demonstrates the great truth which is foreshadowed by a careful and comprehensive examination of the gradations of form which they present in their adult state; namely that they are all constructed upon one plan; that they differ, indeed, in the extent to which this plan is modified, but that all these modifications are foreshadowed in the series of conditions through which the skull of any one of the higher Vertebrata passes.
But if these conclusions be correct, the first problem which I proposed to you,–Are all vertebrate skulls constructed upon a common plan?–is solved affirmatively.
We have thus attained to a theory of general expression of the laws of structure of the skull. All vertebrate skulls are originally alike; in all (save Amphioxus ?) the base of the primitive cranium undergoes the mesocephalic flexure, behind which the notochord terminates, while immediately in front of it, the pituitary body is developed; in all, the cartilaginous cranium has primarily the same structure,–a basal plate enveloping the end of the notochord and sending forth three processes, of which one is short and median, while the other two, the lateral tuberculæ, pass on each side of the space, on which the pituitary body rests, and unite in front of it; in all, the mandibular arch is primarily attached behind the level of the pituitary space, and the auditory capsules are enveloped by a cartilaginous mass, continuous with the basal plate between them. The amount of further development to which the primary skull may attain varies, and no distinct ossifications at all may take place in it; but when such ossification does occur, the same bones are developed in similar relations to the primitive cartilaginous skull. But the theory of the skull thus enunciated is not a 'vertebral theory'; one may have a perfectly clear notion of the unity of organisation of all skulls without thinking of vertebræ.
So much for the first problem before us. I now proceed to the second question, which was, you will recollect, Given the existence of a common plan of organization of all vertebrate skulls; is this plan the same as that of a spinal column?
To deal properly with this question, we must know what is the plan of organization of a spinal column, and that can be learnt only by a careful study of its development, as well as of its adult modifications. Indeed, the latter are unintelligible without a knowledge of the former.
It is impossible to form a clear conception of the essential nature of the process of development of a spinal column, or to compare it [579] with that of the skull, unless we analyse very carefully, and distinguish from one another, the successive steps of that process.21
1. The primary changes of form exhibited by the blastoderm in the region of the spinal column, are, in all the Vertebrata whose development has yet been studied, precisely the same. Two ridges, the "laminæ dorsales," bounding a narrow elongated groove, rise up and eventually unite with one another so as to enclose a cavity–the neural canal. External to the junction of the laminæ dorsales with the blastoderm, the latter is converted more or less completely into the "laminæ ventrales," which become incurved, unite, and eventually enclose the visceral cavity.
A transverse section of the embryo in this state shows a very thin and narrow median plate, separating the neural canal above, from the hæmal or visceral canal below, and passing on each side into thickened masses of blastoderm, which give rise to the laminæ dorsales on the one hand, and to the laminæ ventrales on the other.
For convenience of description, I shall term the median plate the "diaphysial plate," and the lateral ridges the "paraphysial thickenings."
2. The primary histological differentiations, which take place in the rudimentary spinal column just described, are the same in all Vertebrata.
A long filament, composed of indifferent tissue, makes its appearance in the middle of the diaphysial plate, and constitutes the notochord, or chorda dorsalis.
Next, the substance of the paraphysial thickenings undergoes a certain change of tissue at regular intervals, so that they acquire a segmented appearance; solid, broad, darker masses of blastema lying opposite one another in each paraphysial thickening, and being separated by clear, narrow interspaces.
These segments are what the Germans term "Urwirbel," or "primitive vertebræ;" a somewhat misleading name, as they are in every way distinct from what are commonly understood under the name of "vertebræ," even if we use that word in its broadest signification. Professor Goodsir's terms of Somatomes for the segments and Metasomatomes for their interspaces, appear to me to be well worthy of adoption as the equivalents of these "Urwirbel."
3. The next step in the development of a vertebral column, is the histological differentiation of the somatomes. Leaving out of consideration the epithelial and other minor tissues, it may be said [580] that each somatome gives rise to (a) epiaxial muscles, (b) a nerve and its ganglion, (c) the blastema for a vertebral centrum and its neural and haemal arches, and (d) possibly hypaxial muscles; while the metasomatome becomes for the greater part of its extent an "intermuscular septum."
It is unnecessary for my present purpose to trace out particularly the development of any of these parts, except the centrum and its arches.
The blastema, which is specially intended for these parts, appears, in a distinct form, first, in the paraphysial thickenings, and then extends inwards above and below, so as gradually to enclose the notochord in a sheath, while, externally, it passes in the posterior half of each somatome, upwards into the neural arches, and downwards into the hæmal arches.
4. In some Vertebrata the spinal column never gets beyond this stage, nor even so far; but for the present it will be well to confine our attention to those which become completely ossified. In these chondrification is the next step. The blastema of the centra and its prolongations becomes converted into cartilage, but not continuously. On the contrary, at points corresponding with the intervals between every pair of metasomatomes, or with the middle of each somatome, the cartilage is replaced by more or less fibrous tissue. As a consequence, the cartilaginous sheath of the notochord is now divided into regular segments, which alternate with the somatomes, so that each metasomatome abuts upon the middle of one of these cartilaginous vertebral centra.
In every centrum it is necessary to distinguish three tracts or regions:–1. A diaphysial region immediately surrounding the notochord. 2. Two paraphysial regions lying in the paraphysial thickenings. The paraphysial regions give rise to the cartilaginous neural and hæmal semi-arcs, which are primitively continuous with them; so that all parts of the vertebra form one connected whole.
The neural semi-arcs eventually unite in the middle line, and ordinarily send a prolongation upwards from their junction. The hæmal semi-arcs also tend to unite below, but in a somewhat different manner.
5. The last step in the development of the vertebra is the differentiation of its various parts from one another, and their final metamorphosis into their adult form. The notochord, which primitively traversed the centra and the intercentra (intervertebral ligaments, synovial membranes, or the like, between the centra), becomes more or less completely obliterated.
[581] The distal, larger part of the hæmal semi-arc is commonly distinguished from its proximal smaller part, by the conversion of its cartilage into osseous or other tissue, and thus the semi-arc becomes separated into a rib and an articular surface or process, for the head of that rib, to which last the term Parapophysis may be conveniently restricted.
In the dorsal vertebræ of many Vertebrata, the neural semi-arc sends out a process, the Diapophysis, which is eventually met by a corresponding outgrowth of the rib, its so-called tubercle, and the two become firmly connected together.
When ossification occurs, it is a very general, if not invariable rule, that an annular deposit around the notochord takes place in the centrum. I term this the Diaphysis of the vertebra. In some fishes a distinct centre of ossification appears in each paraphysial region, and this may be termed the Paraphysis of the vertebra.
In mammals each end of the vertebra ossifies from a distinct point, and constitutes a central Epiphysis of the vertebra; and in many Vertebrata a part of the under surface of a centrum ossifies separately as a distinct Hypophysis. It is another very general, if not invariable rule, that a distinct centre of ossification appears in, or on, each neural semi-arc or Neurapophysis, and passes upwards, into the spine or Metaneurapophysis; downwards, to unite sooner or later with the diaphysis, or diaphysis and paraphysis; and outwards into the diapophysis.
It is doubtful whether the paraphysis appears as a distinct osseous element in any Vertebrata above the class of fishes, in very few of which even, is it distinguishable in the adult state. Consequently in the higher Vertebrata the paraphysial region is ossified, either from the diaphysis or from the neurapophysis, or from both; and a suture exists for a longer or shorter time at the point of junction of the neural and central ossifications. I will term this the Neurocentral suture. Its position is no certain or constant indication of the nature of the parts above or below it, for it may vary in the same vertebral column from the base of the neurapophysis, to the junction of the paraphysial with the diaphysial region of the centrum.
The number of the centres of ossification in each distal portion of the hæmal semi-arc may vary greatly; the uppermost is called a Pleurapophysis, the lower, Hæmapophyses and Met-hæmapophyses.
Besides these primary centres of ossification of a vertebra, there are others of less constancy. Thus the ends of the metaneurapophyses, diapophyses, and zygapophyses in many Mammalia are ossified from distinct centres; and in the caudal region of many of the higher [581] Vertebrata, outgrowths of the centra unite below to enclose the caudal vessels, and ossify as distinct apophyses.
If the development of the skull be now compared with that of the spinal column, it is found that (1) the very earliest changes undergone by the blastoderm in each are almost identical. The primitive groove extends to the extremity of the future cranial cavity; its lateral walls are continuous with the laminæ dorsales, and these pass into laminæ ventrales, also continuous with those of the spinal region. The laminæ dorsales of the head become the cranial walls and enclose the cerebrum–the continuation of the myelon; the laminæ ventrales give rise to the boundaries of the future buccal and pharyngeal cavities.
2. But at this point the identity of the skull with the spinal column ceases, and the very earliest steps in histological differentiation exhibit the fundamental differences between the two. For, in the first place, in no instance save the Amphioxus, has the notochord as yet been traced through the whole of the floor of the cranial cavity. In no other embryo has it been yet seen to extend beyond the middle vesicle of the cerebrum, or in other words, beyond the level of the rudiment of the infundibulum and pituitary body.
In the second place, the division into somatomes, in all known vertebrate embryos, stops short at the posterior boundary of the skull, and no trace of such segmentation has yet been observed in the head itself
3. Apparently as a consequence of these fundamental differences, the further course of the development of the skull is in many respects very different from that of a vertebral column. Chondrification takes place continuously on each side of the notochord, and beyond it, the two trabeculæ cranii, unlike anything in the spinal column, extend along the base of the cranium. No distinct cartilaginous centra, and consequently no intercentra, are ever developed. The occipital arch is developed in a manner remotely similar to that in which the neurapophysial processes are formed; but the walls of the auditory capsules, which lie in front of them, and which give rise to some of the parts, most confidently regarded as neurapophyses by the advocates of the current vertebral theories of the skull, are utterly unlike neurapophyses in their origin.
So, if we seek for hæmal semi-arcs, we find something very like them, arising from the substance of the basis cranii beneath the auditory cartilage; but there is none connected with the occipital cartilage, and none with the rudiment of the alisphenoid. The palatopterygoid cartilage might be regarded as the haemal semi-arc [583] of the presphenoidal region, though the grounds for so doing are not very strong; but the premaxillary cartilage is something quite without parallel in the spinal column.
4. The mode of ossification of the skull, and the ultimate arrangement of its distinct bony elements, are at once curiously like, and singularly unlike those presented by the spinal column. The basioccipital is ossified precisely after the manner of a vertebral centrum. Bony matter is deposited around the notochord, and gradually extends through the substance of the cartilaginous rudiment of the part.
The combined basi- and pre-sphenoid in Pisces and Amphibia is an ossific deposit, which takes place on the under surface of the basal cartilage in front of the basioccipital, and extends thence completely beneath the pituitary interspace as far as the ethmoid. It might be paralleled by the subchordal ossification in the coccyx of the frog, or by the cortical ossification of the atlas in many higher Vertebrata, if it really underlay a portion of the notochord; but at the very utmost the notochord only extends into its posterior extremity.
In some of the higher Vertebrata, as the snake, the osseous basisphenoid arises in the substance of its cartilaginous rudiment, while the osseous presphenoid underlies its cartilage. In others, both bones appear to arise directly in their cartilaginous forerunners. But nothing can be more irregular than the mode of ossification of the presphenoid, ethmoid and vomer in the vertebrate series, or less like the very constant and regular course of ossification of true vertebral centra.
With respect to the ossification of the lateral and superior constituents of the skull, the development of the exoccipital and supraoccipital does, without doubt, present a very close analogy to that of the separate pieces of the neural arch of some vertebrae in, e.g., a crocodile. The alisphenoids and orbitosphenoids follow in the train of the exoccipitals; but I know not where in the spinal column we are to find a parallel for the double parietals and frontals. But waiving this difficulty, and supposing, for the sake of argument, as was supposed by Oken, that the basisphenoid, alisphenoid and parietals, the presphenoid, orbitosphenoids, and frontals represent the elements of two vertebral centra and neural arches, what is to be made of the petrous and mastoid bones?
The difficulty has been eluded by terming the petrosal a "sense-capsule," the mastoid a "parapophysis." But I apprehend that neither of these explanations can be received for a moment by those who are acquainted with the development of the skull, or with the true homologues of the bones in question in the vertebrate series, or [584] who think that scientific terms should always possess a well-defined and single meaning.
What, in fact, is the origin of the petrous and mastoid bones? There is much reason for believing (according to Remak's late observations) that the membranous labyrinth is primarily an involution of the sensory or epidermic layer of the blastoderm; but however this may be, it is quite certain that the auditory organ is, primarily, altogether independent of the walls of the skull, and that it may be detached without causing any lesion of them, in young embryos.
It is also quite certain that this membranous labyrinth becomes invested by a coat of cartilage, continuous with the cranial wall; but I do not know that there is evidence, at present, to enable one to say positively, whether this cartilaginous auditory capsule is formed independently around the labyrinth, and then unites with the cranium; or whether it is an outgrowth from the cranial walls, which invests and encloses the labyrinth. If the latter be the case, a consistent vertebral theory of the skull must account for all the bones developed out of the auditory capsule; if the former, it must exclude them all, as parts of an extra-vertebral sensory skeleton.
Now the bones developed in the capsule are, in front, the petrosal; behind, the mastoid; above, the epiotic. The first-named bone is admitted, by the most zealous advocates of the vertebral theory, to be a neurapophysis, in all oviparous Vertebrata. Hence they are also bound to admit that, for three centra below and three neural spines bounding the cranial cavity above, there are four pairs of neural arches. More than this, I do not see how it is to be denied that the true mastoid is the morphological equivalent of the petrosal; and in that case there would be five neurapophyses to three central and three neural spines. Furthermore, it is precisely to these two superfluous elements that the only two clear and obvious hæmal arches, the mandibular and hyoid, are attached.
I confess I do not perceive how it is possible, fairly and consistently, to reconcile these facts with any existing theory of the vertebrate composition of the skull, except by drawing ad libitum upon the Deus ex machina of the speculator,–imaginary "confluences," "connations," "irrelative repetitions," and shiftings of position–by whose skilful application it would not be difficult to devise half a dozen very pretty vertebral theories, all equally true, in the course of a summer's day.
Those who, like myself, are unable to see the propriety and advantage of introducing into science any ideal conception, which is other than the simplest possible generalized expression of observed [585] facts, and who view with extreme aversion, any attempt to introduce the phraseology and mode of thought of an obsolete and scholastic realism into biology, will, I think, agree with me, not only in the negative conclusion, that the doctrine of the vertebral composition of the skull is not proven, but in the positive belief, that the relation of the skull to the spinal column is quite different from that of one part of the vertebral column to another.
The fallacy involved in the vertebral theory of the skull is like that which, before Von Bär, infested our notions of the relations between fishes and mammals. The mammal was imagined to be a modified fish, whereas, in truth, fish and mammal start from a common point, and each follows its own road thence. So I conceive what the facts teach us is this:–the spinal column and the skull start from the same primitive condition–a common central plate with its laming dorsales and ventrales–whence they immediately begin to diverge.
The spinal column in all cases becomes segmented into its somatomes; and, in the great majority of cases, distinct centra and intercentra are developed, enclosing the notochord more or less completely.
The cranium never becomes segmented into somatomes; distinct centra and intercentra, like those of the spinal column, are never developed in it. Much of the basis cranii lies beyond the notochord.
In the process of ossificatlon there is a certain analogy between the spinal column and the cranium, but that analogy becomes weaker and weaker as we proceed towards the anterior end of the skull.
Thus it may be right to say, that there is a primitive identity of structure between the spinal or vertebral column and the skull; but it is no more true that the adult skull is a modified vertebral column, than it would be, to affirm that the vertebral column is a modified skull.22
While firmly entertaining this belief, however, I by no means wish to deny the interest and importance of inquiries into the analogies which obtain between the segments, which enter into the composition of the ossified cranium, and the vertebræ of an ossified spinal column. But all such inquiries must start with the recognition of the fundamental truths furnished by the study of development, which, as our knowledge at present stands, appear to me to be summed up in the following propositions:–
1. The notochord of the vertebrate embryo ends in that region of the basis cranii which ultimately lies behind the centre of the basisphenoid bone.
[586] 2. The basis cranii is never segmented.
3. The lamina perpendicularis of the ethmoid has the same morphological value as the presphenoid.
4. The petrosal has the same morphological value as the mastoid, if one is not an integral part of the skull, neither is the other.
5. The nasal bones are not neurapophyses.
6. The branchial arches have the same morphological value as the hyoid, and the latter as the mandibular arc.
7. The mandibular arc is primitively attached behind the point of exit from the skull, of the third division of the fifth nerve.
8. The premaxilla is originally totally distinct from the palatomaxillary arcade.
9. The pectoral arch is originally totally distinct from the skull.
Starting on this basis, it might not be difficult to show that the perfectly ossified skull is divisible into a series of segments, whose analogy with vertebræ is closer the nearer they lie to the occipital region; but the relation is an analogy and not an affinity, and these cephalic sclerotomes are not vertebræ.
1 There is a wide difference, too, in the relative importance of either question to the student of comparative anatomy. Unless it can be shown that a general identity of construction pervades the multiform varieties of vertebrate skulls, a concise, uniform, anal consistent nomenclature becomes an impossibility, and the anatomist loses at one blow the most important of aids to memory, and the most influential of stimulants to research. The second question, on the other hand, though highly interesting, might be settled either way or the other without exerting any very important influence on the practice of comparative Anatomy. 2 The titles of these works are,–Reichert, 'De Embryonum arcubus sic dictis Brachiatlibus,' 1836, which I have not seen; the same writer's essay, 'Ueber die Visceralbogen der Werbelthiere im Allegemeinen,' Müller's Archiv, 1837. Hallmann, 'Die vergleichende Osteologic des Schläfenbeins,' 1837. Rathke, 'Entwickelungsgeschichte der Natter,' 1839. I regret that, in spite of all efforts, I have hitherto been unable to procure a copy of another very important work of Rathke's, the 'Programm,' contained in the 'Vieter Bericht von den naturwissenschaftlichen Seminär zu Koningsberg.' 3 In speaking of these bones I shall avail myself, for the most part, of the useful translation of the Cuvierian nomenclature adopted by Prof. Owen. It is, doubtless, more convenient to say "alisphenoid" than "grande aile" or "aile sphenoidale," and "orbitosphenoid" instead of "aile orbitaire," the slightness of the real change thereby effected being one of its principal recommendations. The adoption of the terms will, of course, not be held to imply any recognition of the justice of the views of either their inventor or their adopter. 4 'Berichte von der Königlichen Zool. Anstalt zu Würzburg,' 1849, p. 40. 6 My reasons for considering this osseous element to be distinct from the supraoccipital will be given below. 8 Though the pterygoid comes close to it in Monotremata. 11 Reichert, however, had already clearly declared this important homology in his 'Entwickelungsgeschichte des Kopfes,' p. 195. 12 Compare Kölliker's account of the primordial skull of a young turtle in the 'Bericht von der Konig. Zool. Anstalt zu Würzburg,' 1849. 13 See for the manner in which this is brought about, Rathke's 'Entwick. d. Natter.' Rathke, it should be said, regards this bone as the tympanicum, but its primitive place and mode of origin are those of the squamosal of the mammal. 14 See Cuvier, 'Ossemens Fossiles,' x. p. 65; and Stannius, 'Zootomie der Amphibien,' p. 68. 15 In adopting the universally known Cuvierian appellations, I merely desire to avoid for the present all theoretical suggestions. 16 See this result, well worked out, by the method of gradation only, by Kostlin (l.c. pp. 328-332), who draws particular attention to the resemblance between the suspensorium of the Amphibia and that of fishes of the Eel-tribe. 17 In Muræna Helena the suspensorium forms an obtuse angle with the axis of the skull, though not so obtuse as in the frog. A strong ligament connects the outer side of the distal end of the quadratum with the maxillary bone, passing outside the lower jaw. If the posterior end of the ligament were ossified, it would correspond very nearly with the "quadratojugal" of the frog 18 Of course in a morphological sense. Whether they are more or less distant in actual space, is not the question. 19 See Note IV., on the suspensorium in fishes. 20 See Note V. for the development of the skull in other Vertebrata. 21 See Note VI. for the details of the development of the spinal column in Vertebrata generally. 22 I feel sure that I met with this phrase somewhere, but I cannot recollect its author.
NOTES.
I.–On the Mastoid in Birds.
The true mastoid of the bird seems hitherto to have escaped notice.
Hallmann says (l. c. p. 33), "In the disarticulated skulls of chickens, I examined the share taken by the different bones in the formation of the labyrinth, by introducing bristles into the semicircular canals, and I found in the proper petrosum (into which the facial and acoustic nerves enter, and which contains the cochlea) the anterior crus of the anterior canal (I term the upper one thus for ready comparison with reptiles) and of the external canal; in the supraoccipital, the upper (=posterior) crus of the anterior canal, and the upper end of the posterior canal; and in the exoccipital, the lower crus of the posterior, and the posterior of the external canal. In other words, the distribution of the canals is as in the scaly Amphibia. For the rest, in birds as in mammals, and probably in all Vertebrata, the membranous semicircular canals are formed connectedly in the cartilage, and the bony parts only gradually invest them. Hence, when the chick's skull is too young, but very little of the posterior canal is to be found in the supraoccipital, which in fact contains somewhat less of the posterior canal than of the anterior and thereby departs from reptiles and approximates mammals.
"At a certain period also, an interval filled with cartilage, through which the semicircular canals shine, is found in the bird's skull between the supraoccipital, the parietal, the squama temporis, and the exoccipital. I see this clearly in the skull of a young Dicholophus cristatus (No. 5605, B.M.). In the skeleton of a young Colymbus cristatus (No. 7172, B.M.), I find that this interval is, on the right side, almost filled up by a small bony plate, which has not as yet combined [587] with the surrounding bones. This appears to me to be a separate pars mastoidea,. which however combines very early with the exoccipital. In the skull of a young goose (fig. 3) (No. 3507, B.M.), this distinct piece (e between s, r, t and l) is still better shown. Subsequently it is altogether indistinguishable from the exoccipital."
I have endeavoured to show, however, that the true mastoid of the bird is to be sought elsewhere; and at any rate the bone described by Hallmann has not those relations which he himself considers essential for a mastoid. It appears to me, that the distinct ossification he mentions is the epiotic bone, which has not yet combined with the surrounding parts.
II.–On the influence of the share taken by the squamosal in the Vertebrate Skull.
In discussing the homologies of the bones of the skull of the crocodile, Cuvier ('Ossemens Fossiles,' t. ix. p. 163) states that "the squamosal and zygomatic bone becomes more and more excluded from the cranium as we descend in the scale of quadrupeds, so that in Ruminants it is rather stuck upon the skull than enters into the composition of its walls;" and it is by this argument mainly that the great anatomist justifies his identification of the quadratojugal of the crocodile with the squamosal, or rather with the zygomatic portion of that bone in mammals (l. c. p. 171).
Professor Owen ('Principes d'Ostéologie Comparée,' p. 55) adopts Cuvier's argument, and pushes it further, endeavouring to show that the disappearance of the squamosal, and as he supposes of the petrosal, from the interior of the skull in Reptilia is sufficient to account for that retrogression of the alisphenoid behind the exit of the fifth nerve, which is the necessary consequence of his identification of the true petrosal with the alisphenoid.
It seems strange that Cuvier should have advanced so weak an argument as that which I have cited; for assuredly Ruminants are not very low in the mammalian scale, nor are they those mammals which most nearly approach reptiles or birds. We must seek these among rodents and monotremes, in both of which the squamosal enters largely into the composition of the cranial walls.
This is particularly the case in that especially reptilian mammal the Echidna. As to birds, it can still less be said that their squamosal disappears from the interior of the skull. Kostlin says on this point ('Bau des knöchernen Kopfes,' p. 206), "The squamosal contributes a small surface to the ridge, which separates the anterior cranial fossa from the middle one. It is here applied above against the parietal, anteriorly against the anterior, and posteriorly against the posterior sphenoidal ala***l and seems in all birds to appear at this point in the cavity of the skull. In the goose its extent is far smaller than in the fowl. The actual size of the ala temporis, however, surpasses that of its inner surface by a great deal. In this respect birds are analogous to the Cheiroptera, Insectivora, and a few Marsupialia, where only a small portion of the squama temporis projects into the cranial cavity. Still more do they resemble the seals, in which this part is entirely enclosed by the parietal and ala temporis, and so is completely separated from the petrosal." Kostlin is in error, however, in assuming that the squamosal is visible in the interior of the skull of all birds, for as we have seen above, such is not the case in the ostrich.
The struthious skull then affords an important test of the value of Professor Owen's argument. If, as he supposes, the disappearance of the squamosal from [588] the interior of the skull causes the alisphenoid to pass behind the exit of the trigeminal, this retrogression ought to have taken place in the ostrich. Nothing of the kind has occurred, however, the trigeminal foramen being a 'trou de conjugaison' between the alisphenoid and the petrosal. It does not even traverse the middle of the alisphenoid, as in the sheep.
It is unnecessary to discuss the effect of the disappearance of the petrosal, as I have endeavoured to prove that it does not disappear in the lower Vertebrata.
III–Connexions of the tympanic membrane in Birds.
According to Kostlin (l. c. p. 216), the tympanic membrane of birds is stretched upon a fibrocartilaginous frame, which is ordinarily attached to the squamosal, exoccipital, basisphenoid, and quadrature. In many gallinaceous birds this frame does not come into contact with the quadrature at all. From these circumstances, and from the fact that the quadratum of birds articulates with the lower jaw and the jugal arch, which is never the case with the tympanic of mammals, Kostlin concludes, with great justice, that the quadrature is not the homologue of the mammalian tympanic.
IV.–On the modifications of the palatosuspensorial arch in Fishes.
I have very briefly stated my views on this subject in the Quarterly Journal of Microscopical Science for October 1858, hoping at that time to enter more largely upon the subject in this place. But the present Lecture and its notes already occupy so much space, that I must reserve a full statement of what I have to say respecting the palatosuspensorial apparatus of fishes for a future occasion.
V.–On the development of the Cranium.
In confirmation of the views which I have adopted, as to the primary uniformity of plan of all Vertebrate crania, I subjoin an abstract of Rathke's most valuable account of the development of the skull in Coluber matrix,2 which contains much incidental information relating to the development of the skull in Vertebrata in general. Vogt's observations on Coregonus and Alytes, and my own on Gasterosteus, Rana and Triton, are in entire accordance with those of Rathke, so far as the primitive structure of the basis cranii is concerned.
The differences between the basis of the skull and the vertebral column in the earliest embryonic conditions are,–
1. That round that part of the chorda which belongs to the head, more of the blastema, that is to be applied, in the spinal column, to the formation of the vertebræ and their different ligaments, is aggregated than around the rest of its extent, and–
2. That this mass grows out beyond the chorda to form the cranial trabeculæ.
The lateral trabeculæ at their first appearance formed two narrow and not very thick bands, which consisted of the same gelatinous substance as that which constituted the whole investment of the chorda, and were not sharply defined from the substance which lay between them and at their sides, but seemed only to be two thickened and somewhat more solid, or denser, parts of that half of the basis of the cranium which lies under the anterior cerebral vesicle.
Posteriorly, at their origin, they were separated by only a small interval equivalent to the breadth of the median trabecula, and thence swept in an arch to [589] about the middle of their length, separating as they passed forwards; afterwards they converged, so that, at their extremities, they were separated by a very small space, or even came into contact. Altogether they formed, as it were, two horns, into which the investing mass of the chorda was continued forwards. The: elongated space between them, moderately wide in the middle, was occupied by a layer of softer formative substance, which was very thin posteriorly, but somewhat thicker anteriorly. Upon this layer rested the infundibulum; and in front of it, partly on this layer, partly on the trabeculæ, that division of the brain whence the optic nerves proceed, and further forwards the hemispheres of the cerebrum. Anteriorly, both traberculæ reached as far as the anterior end of the head, and here bent slightly upwards, so that they projected a little into the frontal wall of the head, their ends lying in front of the cerebrum. Almost at the end of each horn, however, I saw a small process, its immediate prolongation, pass outwards and form, as it were, the nucleus for a small lateral projection of the nasal process of the frontal wall.
The middle trabecula grows, with the brain, further and further into the cranial cavity, and as the dura mater begins to be now distinguishable, it becomes more readily obvious than before, that the middle trabecula raises up a transverse fold of it, which traverses the cranial cavity transversely.3 The fold itself passes laterally into the cranial wall; it is highest in the middle, where it encloses the median trabecula, and becomes lower externally, where it forms, as it were, a short ala proceeding from the trabecula. With increasing elongation, the trabecula becomes broader and broader towards its free end, and, for a short time, its thickness increases. After this, however, it gradually becomes thinner, without any change in its tissue, till, at the end of the second period, it is only a thin lamella, and after a short time (in the third period) entirely disappears.
In mammals, birds, and lizards, that is, in those animals in general, in which the middle cerebral vesicle is very strongly bent up and forms a protuberance, while the base of the brain exhibits a deep fold between the infundibulum and the posterior cerebral vesicle, a similar part to this median trabecula of the skull is found.
In these animals, also, at a certain very early period of embryonic life, it elevates a fold of the dura mater which passes from one future petrous bone to the other, and after a certain time projects strongly into the cranial cavity. Somewhat later, however, it diminishes in height and thickness, as I have especially observed in embryos of the pig and fowl, until at last it disappears entirely in these higher animals also, the two layers of the fold which it had raised up. coming into contact. When this has happened, the fold diminishes in height and eventually vanishes, almost completely.
The two lateral trabeculæ, which in the snake help to form the anterior halt of the basis of the skull, attain a greater solidity in the second period, acquire a greater distinctness from the surrounding parts, and assume a more determinate form, becoming, in fact, filiform, so that the further forward, the thinner they appear. They increase only very little in thickness, but far more in length, during the growth of the head. Altogether anteriorly, they coalesce with one another, forming a part which lies between the two olfactory organs and constitutes a septum. As soon as these organs increase markedly in size, this part is moderately elongated and thickened, without however becoming so dense as the hinder, longer part of the trabeculæ. The prolongations into the lateral projections [590] of the nasal processes, which now proceed from the coalesced part in question, also become but little denser in texture for the present, though they elongate considerably.
The lateral parts and the upper wall of the cranium, with the exception of the auditory capsules or of the subsequent bony labyrinth, remain merely membranous up to the end of the second period, consisting in fact only of the cutaneous covering, the dura mater, and a little interposed blastema, which is hardly perceptible in the upper part, but increases in the lateral walls, towards the base of the skull.
The chorda vertebralis reaches, in very young embryos of the snake, to between the auditory capsules, and further than this point it can be traced neither in the snake nor in other Vertebrata, at any period of life, as manifold investigations, conducted with especial reference to this point, have convinced me.
At the beginning of the third period, the basal plate chondrifies, at first leaving the space beneath the middle of the cerebellum membranous; but this also eventually chondrifies, and is distinguished from the rest of the skull only by its thinness.
Lateral processes grow out from the basal cartilage just in front of the occipital foramen, and eventually almost meet above. They are the exoccipitals.
The two lateral trabeculæ, parts which I have also seen on frogs, lizards, birds, and mammals, chondrify at the beginning of the third period. At first, they pass, separate from one another throughout their whole length, as far as the frontal wall, on entering which they come into contact; are more separate posteriorly than anteriorly, and present, in their mutual position and form, some similarity with the sides of a lyre. But as the eyes increase, become rounder, and project, opposite the middle of the trabeculæ, downwards towards the oral cavity, the latter are more and more pressed together, so that even in the third period they come to be almost parallel for the greater part of their length. Anteriorly, however, where they were already, at an earlier period, nearest to one another, they are also pressed together by the olfactory organs (which have developed at their sides to a considerable size), to such a degree, that they come into contact for a great distance and then completely coalesce; they are now most remote posteriorly, where the pituitary body has passed between them,4 so that they seem still to embrace it. Anteriorly, between the most anterior regions of the two nasal cavities, they diverge from their coalesced part as two very short, thin, processes or cornua, directed upwards, and simply bent outwards.
It has been seen above that the median trabecula does not chondrify, but eventually disappears; in its place, a truly cartilaginous short thick band grows into the fold of dura mater from the cartilaginous basal plate."Where the pituitary gland lies, there remains between the lateral trabeculæ of the skull a considerable gap, which is only closed by the mucous membrane of the mouth and the dura mater. But there arises in front of this gap, between the two traberculæ, as far as the point where they have already coalesced, a very narrow, moderately thick, and anteriorly pointed streak of blastema, which, shortly before the end of the third period, acquires a cartilaginous character, and subsequently becomes the body of the presphenoid.5
[591]"Altogether anteriorly, however, where the two trabeculæ have coalesced, there grows out of this part, from the two cornua in which it ends, a pair of very delicate cartilaginous plates. At the end of the third period both plates acquire a not inconsiderable size, take the form of two irregularly formed triangles, and are moderately convex above, concave below, so as to be on the whole, shell-shaped. The nasal bones are developed upon these, while below them are the nasal cavities, and the nasal glands with their bony capsules.
"The alæ or lateral parts of the two sphenoids do not grow like the lateral parts of the occipital bone out of the basis cranii, whose foundation is formed by the cephalic part of the chorda, but are formed separately from it, although close to it, in the, until then, membranous part of the walls of the cranium.
"The alæ of the presphenoid (orbitosphenoids), which are observable not very long before the termination of the third period, appear as two truly cartilaginous (though they never redden), irregular, oblong, plates of moderate thickness, lie in front of the optic foramina, at the sides of the lateral trabeculæ of the skull, ascend from them upwards and outwards, and are somewhat convex on the side turned to the brain, somewhat concave on the other. The ala magnæ (alisphenoids) are perceptible a little earlier than these. They are formed between the eye and the ear, and also originally consist of a colourless cartilaginous substance; they appear at the end of the third period as irregular four-sided plates, lie at both sides of the anterior half of the investing plate of the chorda, ascend less abruptly than the alæ orbitales, and are convex externally, internally concave.
"The upper posterior angle of each elongates, very early, into a process, which grows for a certain distance backwards, along the upper edge of the auditory capsule, and applies itself closely thereto.
"The auditory capsules, or the future petrous bones,6 chondrify, as it would appear, the earliest of all parts of the skull: the fenestra ovalis arises in them by resorption.
"The ossification of the snake's skull commences in the basioccipital, or at any rate, this is one of the first parts to ossify. At a little distance from the occipital foramen, there arises a very small semilunar bony plate, whose concave edge or excavation is directed forwards; thereupon the bony substance shoots from this edge further and further forwards, until at length the bony plate has the form of the ace of hearts. Its base borders the fontanelle in the base of the skull, which lies under the anterior half of the third cerebral vesicle, while its point is contiguous to the occipital foramen; for the most part it is very thin, and only its axis (and next to this its whole posterior margin) is distinguished by a greater thickness. The cephalic part of the chorda can be recognized in the axis of this bony plate up to the following period. It passes from the posterior to the anterior end of the bony plate, where it is lost, and is so invested by the bony substance of the plate, that a smaller portion of the latter lies on the upper side of the chorda, a larger portion beneath it. On this account it forms, on the upper side of the plate, a longitudinal ridge, which subsequently becomes imperceptible by the aggregation of matter at the sides. On one occasion, however, I saw, in an embryo which was almost at term, a similarly formed and sized bony cone, which, through almost its entire length, appeared merely to lie on the body of the basioccipital, since it had only coalesced with it below."
The nucleus and sheath of the cephalic part of the chorda become gradually broken up and the last trace of them eradicated, as the ossification of the basi[591]occipital proceeds, like the nucleus and sheath of the rest of the chorda wherever a vertebral body is developed.7
The articular condyle is not yet formed. The exoccipitals ossify through their whole length and breadth.
The body of the basisphenoid is formed between the above-mentioned posterior fontanelle of the basis cranii and the pituitary space, 'therefore far from the cephalic part of the chordal. It ossifies by two lateral centres, each of which forms a ring round the carotid canal. The alisphenoids ossify in their whole length and breadth; the orbitosphenoid only slightly, and the presphenoid not at all. The premaxillary bone arises as an azygos triangular cartilage between the cornua of the anterior ethmovomerine plate. It ossifies from a single centre.
"The auditory capsule, or the future petrosal bone, may, even at the end of this period, be readily separated from the other part of the cranial wall, and still consists for the most part of cartilage. On the other hand, the triangular form, which it had before, is not inconsiderably altered, since it greatly elongates forwards, and thus, as it were, thrusts its anterior angle further and further forwards, and becomes more unequal-sided. At the lower edge, or the longer side of it, about opposite to the upper angle, at the beginning of this (third) period, or indeed somewhat earlier, a diverticulum of the auditory capsule begins to be formed (the rudimentary cochlea), and developes into a moderately long, blunt, and hollow appendage, whose end is directed downwards, inwards and backwards and also consists of cartilage. Close above, and somewhat behind this appendage, however, there appears, at about the same time, a small rounded depression, in which the upper end of the auditory ossicle eventually rests; and somewhat later, an opening appears in this depression which corresponds with the fenestra rotunda of man. Very much later, namely, towards the end of this period, the auditory capsule begins to ossify. Ossification commences in a thin and moderately long, hook-like process, which is sent forwards and inwards from the lower hollow diverticulum of the cartilage, and unites with the basisphenoid. From this point it passes upwards and backwards, and, for the present, extends so far that, at the end of this period, besides that process, the diverticulum in question and about the anterior third of the auditory capsule itself, are ossified. Later than at the point indicated, an ossific centre appears at the posterior edge of the auditory capsule, where it abuts against the supra- and ex-occipitals, but extends from hence by no means so far forward as to meet that from the other point. The middle, larger part of the auditory capsule, therefore, for the presents remains cartilaginous.
"In the beginning of the fourth period, a third ossific centre arises in the upper angle of the capsule, whereupon all three grow towards one another. But the mode of enlargement and coalescence of these bony nuclei is very remarkable. They do not unite with one another in such a manner as to form a continuous bony capsule for the membranous part of the labyrinth, but are permanently separated by cartilagino-membranous and very narrow symphyses. On the other hand, one coalesces, in the most intimate manner, with that edge of the supraoccipital which is nearest to it, so that even in the more advanced embryos, this bone and it form a moderately long oblong plate, each end of which constitutes a small, tolerably deep, and irregularly formed shell, containing a part of the anterior or upper semicircular canal. The second bony centre becomes an[593]chylosed with the anterior edge of the lateral part of the occipital bone, and also forms a small, irregularly-shaped, but longish scale, which contains the deeper or lower part of the posterior crus of that semicircular canal, and besides this, the lower sac, or representative of the cochlea of the auditory labyrinth. The remaining bony mass of the auditory cartilage, however, includes the greater part of the membranous portion of the labyrinth, and is the largest. The same phenomenon, viz. that the petrosal bone breaks up, as it were, into three pieces, of which two coalesce with the occipital bone, occurs also, according to my observations, in Lacerta agilis, and probably takes place in like manner, if we may conclude from the later condition of the petrous bone to the earlier, in Crocodilia and Chelonia.. A squama temporis and a mastoid are, as I judge, never formed in Ophidia."
Yet what is the osseous mass which eventually coalesces with the exoccipital but the mastoid? I have indicated above what I believe to be the true ophidian squamosal.
VI.–On the development of the Ossified Vertebral Column.
The concise statement of the general nature of this process which I have given above, is based, partly on the observations of Vogt, Rathke, and Remak, and partly on my own. As great misunderstanding seems to me to have prevailed on this head, I have put together in the present note, all the most important evidence I have been able to collect on this highly interesting subject, accompanying it with a running commentary. I have done this the more willingly, as the accounts of the mode of development of vertebræ in general, in our own language, which I have met with, are strangely meagre.
Development of the Spinal Column in Fishes.
1. Blemmius viviparus (Rathke, 'Bildungs- und Entwickelungsgeschichte des Blemmius viviparus.' 1833).
The surface of the notochord hardens, and acquires a fibro-membranous consistence, while its inner substance becomes glassy and transparent, so that the notochord is separated into sheath and contents, as in the lamprey. A segmentation next takes place in the sheath. "At successive intervals it increases, more and more, in density and solidity, acquires in places almost the constitution of cartilage, and there thus arise a great number of successive, very fine, narrow rings, which are connected by much narrower, far less solid, but also far less transparent and more whitish-coloured parts, like sutures."
When this segmentation has commenced, "a number of cartilaginous, very short, thin and rod-like processes, which run in pairs from each member of the vertebral column, where its upper side passes into the two outer ones, appear, pass upwards in the walls surrounding the spinal marrow, and enclose its lower cords. At first, therefore, each pair of processes are separated by a considerable interval throughout their entire length. Subsequently their upper ends approximate (increasing in length, and at the same time accommodating themselves to the curve of the spinal marrow, and bending round it) more and more closely, till they, at last, meet above the spinal marrow, and soon after this has happened, coalesce into an arch.
"Contemporaneously: with these processes, and in the same way, there arise from the vertebral column (though only from its hinder half, or that which constitutes the foundation of the tail) a number of other processes similar in form and structure, which spring from the junction of the under with the lateral faces of the [594] column. These take the opposite direction to the preceding, tend to enclose the great caudal vessels, and unite in pairs into arches, which lie in a series and correspond with the vertebral segments."
From the segments of that part of the vertebral column which lies between the tail and the head, there grow out, in corresponding places to those in which the crura of the inferior arches take their origin from the vertebral segments of the tail, and in the same manner and at the same time, many cartilaginous processes, which attain, however, only a very slight length, and also take a transverse direction. They might be regarded as lateral pieces of the transverse processes of the higher animals; but it is more probable that they correspond with the ribs of other Vertebrata.
"All these processes are connected with the sheath, but not with the core of the notochord."
As development advances, the ring-like segments increase in breadth, length and, thickness; at the same time they become somewhat cartilaginous, and then ossify. Each widens somewhat more at its ends than in the middle, and so appears a little contracted in the centre. It is only after birth that such an internal thickening takes places as to interrupt the cavity of the vertebral centrum.
The sheath of the notochord is originally of one texture throughout, but the smaller portions, which lie between the vertebral centra, assume a fibrous texture, contemporaneously with the appearance of the latter. The included substance of the notochord loses its peculiar dense and elastic character, becomes first gelatinous, then grumous, and finally resembles a thick serum.
The crura of the upper and lower vertebral arches (in the tail) unite in pairs, and their points of union grow out into spinous processes.
The ossification of the processes which arise from the vertebrae commences at the point of junction of the process with the centrum. "A small bony point arises, which appears to belong to both centrum and process, and from whence ossification extends into both. In each vertebral centrum, therefore, as well of the tail as of the trunk, ossification proceeds from different and distant points."
2. Cyprinus blicca (Von Bär, 'Untersuchungen über die Entw. d. Fische.' 1835).
"At the end of the first day the notochord is covered by something which surrounds it like thin plates; these are the developing bodies of vertebræ. It is clearly observable that these bodies of vertebræ are not undivided rings surrounding the notochord, but that they consist of many pieces united by sutures. This condition also is persistent in the sturgeons. The body of the vertebræ therefore, is formed of the coalescence of many pieces, and a lateral suture seems to indicate that these processes are elongations of the previously observed upper and under vertebral arches."
Von Bär imagines that the unconstricted part of the notochord gives rise to the intervertebral ligaments.
From these observations of Rathke and Von Bär, it would appear as if the annular ossifications which surround the notochord arose by the coalescence of ossific centres, primarily developed at the junction of the apophyses with the centra. My own observations on Gasterosteus, however, show, like those of Vogt on Coregonus, that the centra ossify from distinct rings deposited immediately round the notochord; and I am very strongly inclined to believe that the corresponding primary annular diaphyses of the vertebrae in Cyprinus and Blennius have been overlooked.
3. Coregonus palea (Vogt, 'Embryologie des Saumones,' 1842, p. 104, et seq.).–[595] But it is necessary to distinguish carefully between what we call vertebræ in the adult fish, that is to say, those osseous or cartilaginous pieces intended for the support of the whole body, and more particularly of the spinal cord, and such vertebral divisions as we find in embryos. These last are the general fact, the expression of a constant law according to which all the Vertebrata are developed. The vertebræ of adult fishes, on the other hand, are solid rings, whose presence depends on the type peculiar to each species; consequently their form, and the substance of which they are composed, vary in almost every species.
"The vertebral divisions8 appear very early in the Coregonus–almost at the same time as the notochord; and when the dorsal groove begins to close, they are fine lines, caused, as it would appear, by a greater accumulation of embryonic cells, which, like transverse septa, traverse the entire mass as far as the notochord. These divisions extend forwards, as far as the neighbourhood of the auditory vesicles, but there never exists the smallest trace of them in the head itself. At first they are visible only in the middle of the body; by degrees they move forwards, as far as close to the ear, and backwards, towards the tail, as far as it is formed; but they invade its extremity only when it has attained its full length relatively to the body. At first these lines are all straight and perpendicular to the axis of the chorda; but by degrees, and in proportion as development advances, they become oblique and bend, forming an angle whose apex is directed forwards, land corresponds exactly to the median line of the notochord."
They eventually become the intermuscular septa. Vogt goes on to say,–"The typical structure of the Vertebrata, then, consists solely in these rings of separation, which are formed around a notochord, and nowise in the development of a distinct head, or of other solid pieces of the skeleton, such as osseous or cartilaginous vertebrae," illustrating his case by the Amphioxus.
Each osseous vertebra corresponds to two vertebral segments, namely, to the half of that which precedes, and to the half of that which follows a metasomatome; for it is where the latter reaches the notochord, that the centra and arches take their origin. The centra arise as a double ring of cartilage, internal and external to the sheath of the notochord. The intervertebral spaces always correspond with the middle of the interval between two intermuscular septa, each of which is consequently inserted into the middle of a centrum, while the superior and inferior arches are developed in their plane. They are ossified only long after the centra, which arise as bony rings around the notochord. The intervertebral ligaments are formed from the sheath of the notochord.
4. Prof. Owen ('Principes d'Ostéologie Comparée,' 1855, p. 184) affirms that "In osseous fishes the centrum is ordinarily ossified from six points, of which four begin in the bases of the two neurapophyses and of the two parapophyses, but the terminal concave plates of the centrum are ossified separately."
It is not stated on what fish the observations on which this latter assertion is based were made. Prof. Williamson has already shown ('On the Development of the Scales and Bones of Fishes,' Phil. Trans. 1851) that it is inconsistent with the structure of the adult vertebra; it is not supported by any of those writers who have directly observed the development of the vertebræ of Teleostean fishes, and it is negatived by the observations of Vogt just cited and by my own.
Development of the Spinal Column of Batrachia.
1. Anura (Dugès, 'Recherches sur les Batraciens,' 1835).–In the first period the notochord appears to be divided transversely into "rondelles" or vertebræ, but [596] these are not real divisions; they are appearances produced by the intersections of the muscles which surround the notochord, and of the transverse vascular branches which accompany each pair of nerves when it leaves the medulla. [Dugès thus describes the somatomes.]
In the second period cartilaginous processes, adherent to the notochord, appear in pairs and enclose the medulla. There are as many of these processes as vertebræ will in future exist, and two crests even make their appearance, to form the walls of the coccygeal canal. The apophyses are at first little tubercles; they then bifurcate; one branch becomes the transverse process, the other the neurapophysis with its zygapophyses. In B. fuscus, A. obstetricans, punctatus, and Hyla, where these vertebræ ossify, "two clouds" of ossific matter make their appearance in each vertebra, "as distant from one another as they are from the lateral masses or apophyses," and eventually unite above the notochord so as to form a quadrate ossific centre. This quadrate mass enlarges, but remains concave, not only above, but also in front and behind, and especially below, where it forms a semi-canal or groove, in which the notochord is lodged. The groove is gradually filled up, the notochord undergoing a contemporaneous atrophy, and becoming eventually reduced to a mere ligament. The intervertebral masses are formed altogether independently of the notochord.
In Rana, on the other hand, the primitive centre of ossification of the body of the vertebra is a ring completely enclosing the chorda; in other respects the development of the spinal column resembles that just described.
2. Müller ('Vergleichende Anat. d. Myxinoiden,' 1835) remarks, "that the formation of the primitive elements of the skull (which are different from the secondary osseous ones) takes place in the higher animals, constantly in the same way, is much to be doubted, since variations of the fundamental plan obtain in the vertebral column. In many Batrachia, as Cultrips provincialis, and Rana paradoxa, the bodies of the vertebræ arise only out of the upper primitive vertebral elements. I found, indeed, in the larva of Rana paradoxa, on the under part of the circumference of the chorda dorsalis, a cartilaginous band which was especially well developed posteriorly, in front of the ossification of the coccygeal spine, and was continued, thinner, along the under surface of the chorda, for half the length of the future vertebral column. This cartilaginous band had no fellow, but on the contrary, was thickest in the middle. In the caudal part of the chorda it diminished until it gradually disappeared, so that the inferior arches surrounding the caudal vessels were merely fibrous productions of the external sheath of the chorda. But this inferior cartilaginous band on the chorda of the larva of Rana paradoxa disappears in the greater part of the spinal column, and merely a part of it ossifies to become the basilar part of the coccyx, which Dugès was acquainted with, as well as with the two vertebræ of the coccyx above the chorda; the basilar bone is not a body of a vertebra, but coalesces subsequently with the inferior circumference of the coccygeal vertebrae. In these frogs, the coccyx is the only part which arises from both upper and lower vertebral elements; all the other vertebræ arise in Cultripes and Rana, merely from the upper primitive vertebral elements, which in the course of ossification become divided into arches and central portions. It is only the ossifications of the coccyx which, in these frogs completely enclose the chorda, since that part is eventually composed of two pairs of vertebræ, and a long basilar piece, whose sutures are retained even in the adult R. paradoxa " '(p. 130).
3. Alytes.–With respect to the development of the vertebræ in Alytes obstetricans, Vogt ('Entwickelungsgeschichte der Geburtshelferkröte' 1842) states that cartilaginous rings appear in the sheath of the chorda, as rudiments of the [597] centra. Contemporaneously with these the cartilaginous neural arches are developed in the wall of the canal of the medulla; nothing is said as to the mode of ossification.
4. In both Rana temporaria and Triton, I find that the diaphysis of the vertebra arises as a saddle-like patch, upon, and in immediate contact with, the dorsal surface of the notochord; the layer of osseous matter is at first exceedingly thin, and gradually extends round the notochord until in most of the frog's vertebra and in all of those of the Triton, it forms a complete ring. The osseous deposit in the arches is quite distinct, and has, in the frog, the form of a thin bony sheath investing their cartilaginous basis. The diaphysis of the sacral vertebra remains open below long after the others, and after its neural arch is completely ossified.
5. The development of the coccyx of the anourous Batrachia has been well described by Dugès (l. c. p. 108). The two neural arches originally formed in this region ossify and unite above the spinal cord, and at the same time two osseous centra, which very soon coalesce with them, are formed. These centra are incomplete arcs, open below, where they embrace the notochord. A long cartilaginous plate, however, arises on the ventral surface of the notochord, extending backwards far beyond the level of these posterior coccygeal vertebræ. It ossifies, and eventually becomes anchylosed with the bodies of the coccygeal vertebræ to form the coccyx. Such is the substance of Dugès' views, which, as has been seen, have been confirmed in all essential points by Müller.
Prof. Owen, however, gives a very different account of the matter.
"The vertebra of the tail of the larvæ of the Anura are seen distinctly only in the aponeurotic stage. When chondrification occurs, the operation of absorption and coalescence takes place, and two long neurapophyses only are established on each side; the ossification of these plates extends into the fibrillar sheath of the rest of the coccygeal notochord, and when the perishable parts of the tail of the larva have been absorbed, and the fore- and hind-legs are developed, they constitute by their connation the elongated, osseous coccygeal style, often hollow, of the anourous Batrachia." (Principes d'Ostéologie,' 1855, p. 186.)
Prof. Owen does not state on what anourous batrachian his observations were made, nor does he notice the wide discrepancy between his views and those of Dugès. I have carefully studied the development of the coccyx in the common frog, and my observations are in entire agreement with those of Dugès. Nothing can be more clear than the primitive entire independence of the inferior cartilaginous plate, which by its ossification constitutes the major part of the coccygeal style, from the two neurapophyses and the rudimentary diaphyses which correspond with them.
Development of the Spinal Column of Reptilia.
1. Ophidia (Rathke, 'Entw. der Natter,' 1839).–"Quadrate plates of a more solid substance than the rest of the blastema appear on each side of the notochord, in the middle of the body, where they are at first largest, diminishing in size backwards and forwards. At first they extend neither into the dorsal nor into the ventral plates of the embryo.
"These plates increase in length, and those of each pair grow towards one another above and below. Each plate grows out into two branches above and below. The inner branches lie in close contact with the notochord, and coalesce with those of the opposite side, so as to form rings which eventually become the bodies of the vertebra. The upper outer branch extends into the wall of the [598] neural canal, and, eventually uniting with its fellow, forms the neural arch. The lower outer branch extends into the ventral wall and becomes the rib."
The ring grows both externally and internally, so as to constrict the notochord (which softens and acquires a grumous consistence), and then becomes converted into bone, so that the notochord is surrounded by a series of bony rings. The notochord takes no part in the formation of the articular intervertebral surface, which is an apophysis and not an epiphysis; the neural arches ossify much later than the centrum, from a single point in the middle of each. The inferior processes are outgrowths of the substance of the vertebra, and in the caudal region are, from the first, double.
"On each side of the body of the vertebra, where the ribs and the vertebral arches radiate from it, the condensed blastema of which it originally consists, grows out slowly, but considerably, and becomes developed (gradually undergoing chondrification) into a plate, whose largest surfaces are vertical, which increases more in length than in breadth and thickness, and which gradually drives the rib and vertebral arch further away from the axis of the vertebral column. At the end of this period it is almost as thick as the length of the vertebra; it then appears, when viewed from in front or behind, as a short irregular oblong, one of whose angles passes into the body of the vertebra, and one of whose shorter sides is turned outwards and upwards. From this side passes one crus of the vertebral arch, while from the side which is turned outwards and downwards at least in most of the vertebra, a rib is developed, so that these processes are connected with the originally existing part of the body of the vertebra, only mediately, by the plate in question.
"The crura of the vertebral arch ossify from their middle towards both ends, the process commencing very soon after the ossification of the centra; but after ossification has begun in them, these originally filiform parts widen into broad oblong plates, which, in the greater part of the body, come into contact only towards the end of the period, and in the tail and hindermost part of the body, only in the following period.
"The ribs ossify far later, and also from the middle towards the end. Before, however, an ossific centre is developed in them, the cartilage, of which the rib now for the most part consists, becomes articulated with the rest of the vertebra. The plate, lastly,9which forms the union between a rib, a vertebral arch and a vertebral body, and subsequently forms a part of the body of the vertebra, ossifies only in the following period.
"The relation of the ribs to the bodies of the vertebræ, therefore, is originally just the same as that of the crura of the vertebral arches to them: just as one of these crura, does each rib arise as an outward growth of that vertebral body which is the first formed of all these parts; whilst, however, the crura of the vertebral arches are directed upwards in order to enclose the spinal cord, the ribs grow downwards to enclose the viscera of organic life, and in this way their greater length in the snake, and a multitude of other Vertebrata, is explicable.
"The transverse processes of the caudal vertebra exhibit the same relations to the bodies of the vertebræ as the ribs in the dorsal region. They arise in the same places as these; become, in like manner, removed, together with the crura of the arches, by lateral outgrowths of the body from its axis; and, before the ribs are articulated, they pass quite imperceptibly into the processes in question. The transition is the more remarkable, as in the snake the hindermost rib is split, in [599] just the same manner as the transverse processes of the three or four succeeding caudal vertebræ
"If we consider, in the first place, the relation of the parts in the adult snake, we find that the penultimate rib, near its upper or inner end, gives off from its upper side, a small process directed upwards and outwards; however, in the last rib this process is about a quarter as long as the remaining part of the rib, which lies external to and below it, so that the whole bone has the form of a two-pronged hayfork, not yet fastened to a handle, and one of whose prongs is for the most part broken off. The same fundamental form is possessed by the transverse process of the first caudal vertebra; and the difference between it, and the rib which lies immediately before it, lies principally in this, that it is not, like the rib, articulated with its vertebra, that its upper half or prong is almost equal in length to the lower, and that, regarded as a whole, it is not half so long as the hindermost rib. The transverse processes of the succeeding caudal vertebræ have quite the same form as their predecessors, but gradually diminish in length backwards. As to the development of the ribs and transverse processes in question, they, like almost all the other ribs, are originally sent out as quite simple rays from the bodies of their vertebra; very soon, however, there arises on the upper side and neck of the ray where it passes out from the vertebral body, an outgrowth which elongates more or less, also assumes a ray-like form, and has its free end directed outwards. Thus a fork is produced, whose one prong is more or less thicker than the other.
Rathke suggests that the accessory ribs of many fish are probably developed in this way, the upper prong becoming articulated with the lower.
"The two or three anterior subvertebral processes in the tail are, and remain, quite simple, like the similar processes of the cervical and many dorsal vertebræ The two halves of the others, which arose as separate lateral processes, remain permanently distinct.
"All the newly commencing inferior processes arise as paired outgrowths of the bodies of the vertebræ."
"The ribs are not less outgrowths (ausstrahingen) of the bodies of the vertebræ than the crura of the arches, as I can say from my investigations on fishes, snakes, lizards, birds, and mammals; even when the bodies of the vertebral are completely chondrified, the ribs form a connected whole with them, but subsequently they become articulated, and are thereby essentially distinguished from the crura of the vertebral arches. In some animals the articulation takes place and retains close to the bodies; in others it takes place also close to the bodies, yet, afterwards, a process grows out between the rib and the body, by which the rib is more or less thrust out; this is the transverse process; where it occurs, the rib is, in all cases, at first united only with it; sometimes, however, a process grows out from the rib (the so-called head with its neck), by which it becomes immediately attached to the body of the vertebra itself, so that it is doubly united with the body. In many cases the rib may also become articulated at some distance from the body, and thus break up into rib and transverse process."
"As respects the ribs of the higher Vertebrata, together with their transverse processes, the development of the snake teaches us, that although they are subsequently seen to be in close connexion with the crura of the vertebral arches, they grow out, not from the base of these arches, but far from them, out of the bodies of the vertebræ themselves; where they have arisen, however, each lateral half of the body of the vertebra increases in thickness, in such a manner that it acquires an ala, which drives the crus of the vertebral arch and the rib further [600] and further from the axis, until at length it appears as a common trunk for both, and therefore may easily deceive one into supposing, that the rib is given off from the base of the crus of the arch, and is a process from it."
2. Lacertilia (Rathke, 'Ueber die Entwick. d. Schildkröten,' 1848, pp. 65-67).–In the lizards (as in the snakes), the osseous centra of the vertebræ appear as rings, which do not so closely embrace the notochord (as they do in fishes and Batrachia), being separated from it by a layer of cartilage.
3. Chelonia (Rathke, 'Schildkröten,' p. 65-67).–In the Chelonia two bony rings arise, the one on the outer surface of the cartilaginous basis of the centrum, the other close to the notochord; the rings thicken and eventually coalesce; the ossification of the arches takes place quite independently of that of the centrum. The notochord takes no essential part in the formation of the articulations between the vertebræ, but runs like a thread through them.
Development of the Spinal Column of Birds.
In the Chick (Remak, 'Untersuchungen über die Entw. die Wirbelthiere,' 1855).–The upper and middle (sensory and motor) layers of the germ coalesce to form the axial plate (primitive streak); the lateral halves of this plate thicken and leave between them a groove, the primitive groove; immediately below the groove, and parallel with it, the notochord appears in the axis of the motor layer. The sensory layer of the axial plate is now the medullary plate; from it the nervous centre is developed; the motor layer is termed by Remak the 'Urwirbel-platte,'–the primitive vertebral plate.
The primitive vertebræ (Urwirbel, somatomes) first appear in the dorsal part of the embryo, as opaque portions of the substance of the primitive vertebral plates, which extend from the sides of the chorda into the lateral plate, or that thickened part of the motor layer with which the primitive vertebral plates are immediately continuous. These primitive vertebræ are the result of a sort of segmentation of the motor layer; they acquire a cubical form, and are separated by clear, narrow interspaces (metasomatomes).
At the beginning of the third day the ventral surface of each primitive vertebra has an almost square shape with rounded angles; the transverse section is no longer square but three-sided, the upper and outer faces having merged into one convex face; the surface turned towards the medullary canal is four-sided and a little concave.
The inferior internal edge of the primitive vertebra grows out towards the notochord, and having reached its outer side, it divides into two lamellar processes, which, coalescing with those of the other side, surround the notochord and constitute the blastema of the vertebral column.
The dorsal layer of the primitive vertebra becomes converted into muscle, and forms a segment of the dorsal muscles; the anterior portion of its substance, beneath this, becomes the spinal ganglion; the posteriors the rudiment of the neural arch and rib. The latter extends backwards beyond the boundary of its primitive vertebra into the region of the next, so that it appears to be divided by the clear line of separation into a larger anterior and smaller posterior portion.
The axial portion of the vertebral column does not become segmented in correspondence with the divisions between the primitive vertebra, but midway between them, so that the lines of separation between the primitive vertebræ correspond with the centres of the permanent vertebræ, each of which may thus be said to be formed by the coalescence of the posterior half of the axis of one primitive vertebra with the anterior half of the next following.
[601] Rathke, 'Schildkröten,' p. 66.–In birds, the centra commence as bony rings which closely encircle the chorda, and lie internal to the general cartilaginous mass of the vertebra. The bony substance extends inwards and constricts the notochord, outwards it permeates the vertebræ.
"In the caudal vertebræ, and perhaps in all the cervical vertebræ, the bony substance of the rings extends gradually to the, surface of their centra. In the dorsal vertebræ, on the other hand, there is formed at the fifteenth day, independently of these rings, a broad though thin bony plate on the upper, and a second on the lower face of the centrum, with which the substance of the ring, as it extends, coalesces. The notochord at the eighteenth day may be seen traversing the intervertebral articular cavities like a thread."
Development of the Spinal Column of Mammalia.
Rathke, 'Schildkröten,' pp. 66, 67.–"In the pig and sheep the bony substance is deposited immediately around the notochord, in such a manner that, at first, as in birds, it forms a narrow and thin ring, from which it passes partly towards the surface, partly towards the ends of the separate centra, and after a time reaches the surface, but not the ends. To complete the centra, there arise in the latter two special disks of bone for each vertebra, which afterwards apply themselves to the previously ossified middle part, and wholly coalesce with it.
In pig embryos of 1 in. to 1 in. 3 lines in length, the notochord ran straight through already existing rudiments of the intervertebral ligament like a delicate filament" (p. 77).
Bischoff, in his various works, shows that the earliest changes in the vertebral column of mammals are the same as in birds.
"In like manner as in the Mammalia and birds, the ribs in the Chelonia grow out as simple rays from the neural arches (Bogenschenke/n) of the vertebra, quite close to their bodies. Very close to the places where they have arisen, however, the ribs of birds and mammals send, as a rule, a process downwards and inwards, which increases more or less in length and thickness, enlarges somewhat at its free end, and becomes closely applied thereby to one, or two, bodies of vertebræ, by the intermediation of the articular capsule which now becomes formed. This process is the neck and head of the rib."–Rathke, Schidkröten, p. 97.
"The cervical transverse processes of birds and Mammalia attain their forked form in quite a different manner from the ribs, namely, by the coalescence at one end, of what are properly two transverse processes which have grown out of the vertebra, while, on the other hand, a rib has become forked, because, though originally a perfectly simple ray, it has sent out a secondary process from one of its ends."
It results from the observations which have just been detailed, that with certain real or apparent exceptions which have been duly noted, there is a very great uniformity in the mode of development of the vertebral column in all Vertebrata.
The primary processes up to, and inclusive of, segmentation or division into somatomes, appear to be the same in all; and there is every reason to believe that the somatomes become differentiated in the same general way.10 There seems to be no difference, save in degree, in the chondrification which takes place in the immediate neighbourhood of the notochord and in the neural and hæmal [602] arches. The intercentra correspond in fishes, as in Amphibia and birds, to the middle of each somatome; and it does not appear that they are, to any appreciable extent, produced by the metamorphosis of the notochord.
When ossification takes place, the diaphysis appears as a ring, or part of a ring, in immediate contact with, or very close proximity to, the notochord, which it usually embraces completely, though in the rare case of some Amphibia, only partially. The diaphysis then increases inwards so as to constrict the notochord, and outwards, so as to invade the centrum more and more. A distinct ossification is commonly formed in each neural arch, and one or more others in each hæmal arch.
In the higher or abranchiate oviparous Vertebrata there would seem to be no other centres of ossification in the vertebra than the five just mentioned, except those of the terminal epiphyses. In fishes, on the other hand, a distinct centre–which might be termed the paraphysis–is occasionally found in the paraphysial portion of the centrum.
The dorsal vertebræ of a young carp exemplify this structure remarkably well. The diaphysis is represented by an annular osseous ring, which surrounds the notochord and gives off a vertical median process or plate, and two inferolateral plates, which unite the hollow bony cones into which the osseous ring dilates in front and behind. The ossified neurapophyses expand into wedge-like lower ends, which embrace the vertical plate of the diaphysis, and whose apices come into contact with its annular part.
A thick cuneiform mass is interposed between the base of the neurapophysis and the inferolateral plate of the diaphysis on each side. The outer surface forms part of the general contour of the vertebra, and is not produced into a distinct process, though it represents a parapophysis, and gives attachment to the broad head of the distinctly ossified rib. The outer half of the mass is ossified as a distinct paraphysis; the inner in the young carp is still cartilaginous. In the adult the whole wedge-like paraphysial portion of the centrum is ossified; but, instead of becoming united with the diaphysis and neurapophysis, it is anchylosed with the rib, and seems to form its head. In the pike, the paraphysis, more or less produced into a parapophysis, remains distinct from both rib and diaphysis, and the latter occupies a very much larger share of the whole vertebra.
As I have said above, no distinct ossific centre appears to be developed in the paraphysial region of the centrum in any of the abranchiate Vertebrata; but it becomes ossified partly by the encroachment of the neurapophysial, and partly by that of the diaphysial, ossifications.
These two ossifications may coalesce so as to leave no trace of their primitive distinctness, as in Ophidia, Lacertilia, and birds; or as in mammals, Crocodilia, Chelonia, and many extinct reptiles, they may remain for a long time, or permanently, separated by a suture, which may be termed the "neurocentral suture."
It is very commonly assumed that this neurocentral suture is a sort of morphological landmark, and that it always indicates the boundary between the neurapophysis on the one hand, and the diaphysis or osseous centrum on the other; so that any process which is given off from the vertebra above the suture, is supposed to arise from the neurapophysis, while those given off below it only, are said to arise from the centrum.
It is only necessary to cite a few facts, which may be readily verified, however, to show that the neurocentral suture is of no value as a test of the nature of the parts above and below it.
[603] In man and in the pig, the heads of the ribs, whether dorsal or cervical, are (as Retzius has well pointed out) articulated above the neurocentral suture, and therefore, if we accept the ordinary definition, not to the centrum at all, but to the neurapophysis. Furthermore, if we accept the ordinary view, the "inferior transverse processes" in the neck of these animals are not parapophyses, but second diapophyses, inasmuch as they arise from the neurapophyses, and not from the centrum.
The "transverse processes" of the lumbar vertebræ are usually given off above the neurocentral suture, and are therefore called "diapophyses." In a young Dugong, in the Museum of the Royal College of Surgeons, I find that, in the two hinder lumbar vertebræ, these transverse processes are given off below the neurocentral suture.
In the Echidna the head of every rib is attached to the centrum, or below the neurocentral suture; and in the neck, this suture lies between the upper and lower transverse processes.
Thus, if we follow out logically the view that the neurocentral suture indicates the boundary between the neurapophyses and the centrum, and if we accept the current definition of diapophyses and parapophyses, we arrive at the conclusion, that, in the cervical region, man and the pig have vertebræ with two diapophyses and no parapophysis, while the monotreme has a parapophysis and a diapophysis on each side; that, in the dorsal region, the ribs in man and the pig are connected only with the neurapophyses, while in the monotreme they articulate only with the centra; that the transverse processes of the anterior lumbar vertebræ of the dugong are diapophyses, while those of the posterior ones are parapophyses
The crocodile and some extinct reptiles, such as the Ichthyosaurus, whose ribs are throughout attached to the centra, afford still more striking instances of the confusion which would be produced by taking the neurocentral suture as a morphological boundary.
Müller has argued that it is a distinctive character of fishes to have the ribs attached to the centre of the vertebræ or to parapophyses, and his views have been adopted by other anatomists; but the ribs of the Ichthyosaurus and of the Echidna are as completely and solely attached to their centra as those of any fish; so that if we merely take the facts furnished by anatomy, the doctrine that there is anything peculiarly piscine in the attachment of the ribs to the centra only, falls to the ground.
But it may be urged that the connexion of the head of the rib with the centrum, in mammals and the higher Vertebrata, is secondary, that with the diapophysis being the primary and essential one. This is a very widely current doctrine, and it is sanctioned by Rathke, as we have seen in the passages quoted above, though they are not quite consistent with one another. It is with great hesitation that I venture to contravene the distinct statements of so eminent and accurate an embryologist as Rathke, but my own observations lead me to precisely the opposite conclusions.
In the spinal column of embryos of the mouse 7-8ths of an inch long, for instance, I find that the posterior dorsal vertebræ (A. fig. 10) have no diapophyses, and that the ribs have no tubercles, but that their heads pass directly into the cartilaginous substance of the centrum. Further forwards (B, C) both diapophysis and tubercle become more and more developed, until at length they come into contact and articulate in the ordinary way. Finally, in the cervical region (D) the rib and the diapophysis are, even in this early stage, confluent.
These facts are, I conceive, wholly at variance with the supposition that the [604] primary connexion of the ribs is with the diapophyses. On the other hand, they teach that the ribs in the mammal are, as in the fish, primarily continuous with the centre of the vertebræ–a result which is in perfect accordance with the ordinary embryological relations of the higher and lower animals.
The hæmapophysial cartilage passes off from the paraphysial cartilage on the one hand, just as the neurapophysial cartilage is given off from it on the other.
If the hæmapophysial cartilage becomes divided into rib and process, at some little distance from the centrum, the rib is said to be attached to a parapophysis; and I believe that the only consistent definition that can be given of a parapophysis is, that it is developed from the proximal end of a hæmapophysial arch.
Fig.10.–Vertebræ of an embryonic mouse 7/8ths of an inch long. A. The penultimate dorsal vertebra. B. A middle dorsal; and C'. an anterior dorsal vertebra. D. A posterior cervical vertebra. N. Metaneurapophysis or neural spine. N.A. Neurapophyses. Z. Zygapophysis. di. Diapophysis. P. Rib or pleurapophysis. t. Its tubercle. D. Diaphysis. The ossified parts are shaded, the cartilaginous, dotted.
If the paraphysis is merged in the general body of the vertebra, and the rib becomes distinctly articulated close to it, the attachment of the rib is said to be to the centrum.
If the paraphysis is bent upwards, so as to pass insensibly in direction into the neurapophysis, and becomes ossified in continuity with the latter, the head of the rib is said to be attached to the neurapophysis, although, in truth, the head of the rib, or at any rate the proximal end of the hæmapophysial arch of which that rib is a part, always retains, so far as we have any evidence, its primitive connexion with its vertebral centrum, into whatever new ones it may enter.
If the neurocentral suture does not define the lower limits of a neurapophysis, and if the true definition of a parapophysis is that given above, it is obvious that [605]°ur nomenclature of the parts of the dorsal and lumbar vertebra throughout the vertebrate series, requires a thorough revision.
To this subject I hope to return on a future occasion.
VII.–I subjoin the views of Vogt, and the criticisms of the late great anatomist Johannes Müller upon them, as the best means of exhibiting their relation to those I have advocated.
"If we ask ourselves what we mean by vertebra the primary segments of the still indifferent tissue round the chorda, which arise in all vertebrate embryos, are the first things to suggest themselves. These persist only in the lowest grades of vertebrate animals, while in the higher they disappear, in consequence of the more and more complete development of secondary organs, especially of the extremities; so far as we are able to trace these segments, so far is there a formation of vertebra
"But there is at once a difficulty, when we endeavour to find these segments in the rudiment of the skull of any vertebrate embryo. It is true that many inflexions may be observed which appear to correspond with such vertebræ, but unfortunately these do not appear in the same places in different embryos; and besides, these inflexions and curvatures of the base of the skull are not in the least similar to the sharply and clearly defined intervals between the primary vertebræ. The first of these intervals is always formed behind the auditory vesicles, and lies therefore between the occiput and the first cervical vertebra; further forwards, as has been said, no such interval is discoverable. But in the Cyclostome fishes, which represent this embryonic condition, no vertebral divisions of the skull are discernible; in fact we have in the Myxinoids, only the chorda with its sheath and muscular and cutaneous vertebral rings, which are repeated up to the skull, but there cease. The skull of the Myxinoids, like that of the higher cartilaginous fishes, cannot by any amount of violence be forced under the vertebrate type. In the skull, then, the primary vertebral segments are wanting However, they might be obliterated by the early development of the organs of sense, or by the aberrant development of the brain.
"But there remains a second means of discovering the cranial vertebra, by examining the solid cartilaginous and bony basis of the skull; though here also we meet with insuperable difficulties. As the primitive type of the more solid bodies of the vertebræ, we have everywhere cartilaginous rings arising out of the sheath of the chorda, and deposited around its nucleus. Whether they arise as lateral halves or as entire rings, whether they embrace the chorda completely or only above or below, is a matter of no essential moment. But are such cartilaginous rings deposited around the chorda, discoverable in the skull? They will be sought for in vain unless it be in the last, occipital, cranial vertebra; in this we still find all the characters of a vertebra–the investment of the chorda, the chondrification in the sheath of the chorda. But the chorda does not pass into the so-called first and second cranial vertebræ; it invariably ends, as Rathke justly states, between the auditory capsules, and never passes into the body of the second cranial vertebra, let alone that of the first. The lateral cranial trabeculæ, which bear the two anterior cranial vertebræ can by no possibility be regarded as centra of vertebræ since in this case the characteristic feature, the being traversed by the chorda, is entirely absent. Again, these lateral trabeculæ are continued uninterruptedly forwards, below the first division of the brain, showing no trace of a median division. But in what part of the vertebral column has it ever been seen that two vertebræ arise united and afterwards divide?
"It has therefore become my distinct persuasion that the occipital vertebra is [606] indeed a true vertebra, but that everything which lies before it is not fashioned upon the vertebrate type at all, and that all efforts to interpret it in such a way are vain; that therefore, if we accept that vertebra (occipital), which ends the spinal column anteriorly, there are no cranial vertebræ at all."–Vogt, Entw. d. Geburtshelferkröte, pp. 98-100.
"Vogt, and in the present work Agassiz also, contest the justice of the theory of the composition of the skull of several vertebræ, and will only admit an occipital vertebra, because the embryonic chorda, according to Vogt's investigations, extends no further in the skulls of fishes and Amphibia. In this, in my opinion, too much stress is laid upon a single result of embryological investigation. That, however, the chorda in the frog's larva extends beyond the base of the occiput, further than where the slight trace of the basioccipital is ultimately formed, I have myself seen. Even the anterior part of the vertebral column of the Rays shows that the chordal system, out of which, according to my own and Vogt's observations, only the central part of the fishes' vertebra proceeds, may be abortive, whilst the cortical part of the vertebra, which arises in quite a different way, is at its maximum of development. In a longitudinal section of the anterior part of the vertebral column of a Ray, it is seen that the central parts of the vertebra, in the axis of the vertebral column, or those parts which are developed from the chordal sheath alone, become finer and finer anteriorly (although the column still exhibits vertebral divisions), and at last cease entirely, without reaching the anterior end of the vertebral column. On the other hand, Branchiostoma lubricum shows us the opposite extreme; the chorda passes beyond the anterior end of the skull, beyond the mouth and the eyes, far into the extremest end of the snout.
"This remarkable fact, first observed by Sundevall, was very surprising to me, since in consequence of my studies up till that time, I regarded the existence of three vertebræ in the proper cerebral cranium as certain, at least I considered the assumption of a fourth ethmoidal vertebra to be uncertain and undemonstrated.
"For now I saw at once, that it was undoubtedly possible that the cephalic vertebral column might extend further forwards. There need not always be three cranial vertebræ developed in the head; in birds, reptiles, and fishes, the most anterior vertebra is abortive, and is even entirely wanting in some families; but, in the Mammalia and man, three cranial vertebræ are without exception discoverable in the basis cranii, either in the fœtus, or in many cases even in young or middle-aged animals–the occipitale basilare, sphenoideum basilare, posterius and anterius; these also occur in fish. How far the chorda primitively extends in Mammalia is not yet made out; but even although it should not reach through the whole basis cranii, this, from the reasons which have been stated, would be no good argument."–Joh. Müller, Bericht. cclxviii-ix., Müller's Archiv, 1843.
Notes
1 Alisphenoid and petrosal, mihi. 2 'Entwickelungsgeschichte der Natter,' 1839 3 What Rathke terms the 'middle trabecula,' appears to be only very indistinctly developed in Fishes and Amphibia. 4 The pituitary body, however, as Rathke now admits, does not pass between the trabeculæ, and is developed in quite a different manner from that supposed in the memoir on Coluber. 5 Compare with these statements, the figures and descriptions given above of the embryonic cranium in Gasterosteus and Rana. 6 It will be found from Rathke's statements, further on, that the future petrous bone only represents a portion of each auditory capsule. 7 In the stickleback it has appeared to me that the wall of the anterior conical termination, of the notochord in the basis cranii becomes ossified, or at any rate, invested by an inseparable sheath of bony matter, just in the same way as the 'uro style' is developed in the tail. 8 Metasomatomes, their interspaces being the somatomes. 9 Paraphysial cartilage. 10 The relations of the ganglion to the rudiment of the rib and neural arch and segment of the dorsal muscles in the mouse's embryo are the same as in that of the bird.
|
THE
HUXLEY
FILE

|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
{kind=link}