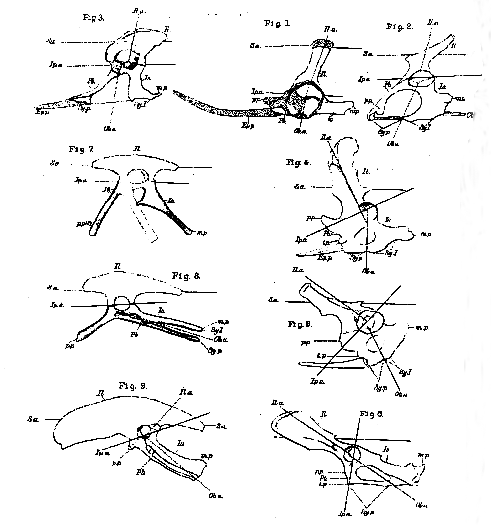
[345] IN the course of the following observations upon the typical characters and the modifications of the pelvis in the Mammalia, it will be convenient to refer to certain straight lines, which may be drawn through anatomically definable regions of the pelvis, as axes. Of these 1 shall term a longitudinal line traversing the centre of the sacral vertebræ, the sacral axis (Plate 8 [Plate 25], S. a.): a second, drawn along the ilium, dorso-ventrally, through the middle of the sacral articulation and the centre of the acetabulum, will be termed the iliac axis (Il. a.); a third, passing through the junctions of the pubis and ischium above and below the obturator foramen, will be the obturator axis (0b. a.); while a fourth, traversing the union of the ilium, in front with the pubis, and behind with the ischium, will be the iliopectineal axis (Ip. a).
The least modified form of mammalian pelvis is to be seen, as might be expected, in the Monotremes, but there is a great difference between Ornithorhynchus and Echidna in this respect, the former being much less characteristically mammalian than the latter.
In Ornithorhynchus (Plate 8 [Plate 25], fig. 4), the ilium is remarkably narrow, and the angle between the iliac and the sacral axis is large, so that the ilium is but very slightly inclined backwards. The iliopectineal axis, nearly at right angles with the iliac axis, is inclined to the sacral axis at an acute angle; while the obturator [346] axis is nearly perpendicular to the sacral axis, and the obturator foramen is relatively small. The front margin of the cotyloid end of the pelvis sends off a very strong pectineal process (p. p.), from the inferior basal part of which a short, obtuse tuberculum pubis (t. p.) projects. Between this and the symphysis, the base of the marsupial bone (Ep. p.) is attached. The ventral rami of the pubes are short and, like those of the ischium, they are united throughout their whole length in a long symphysis, the ischial division of which (Sy. I.) is as long as, if not longer than, the pubic division (Sy. p.). The cotyloid ramus of each ischium gives off a stout elongated metishchial process (m. p.) backwards.
In Echidna (Plate 8 [Plate 25], fig. 5), on the other hand, the ilium is much broader; while the iliac axis inclines downwards and backwards, at an acute angle with the sacral axis. The iliopectineal axis being still at right angles with the iliac axis, makes a much larger angle with the sacral axis; and the obturator axis is inclined from above, at an angle of nearly 45° to the sacral axis, downwards and backwards. In fact, the change in the general character of the pelvis seems to result from its ventral elements having been carried backwards and upwards by the backward and upward shifting of that portion of the ilium which lies below the level of its articulation with the sacrum. There are other changes by which the aspect of the pelvis is much altered. The inner wall of the acetabulum is incompletely ossified, but, in other respects, the pelvis makes a considerable approximation towards the ordinary mammalian form. Thus the pectineal process is represented by a less prominent and more elongated ridge; the metischial process widens out into a mere triangular expansion or "tuberosity," of the ischium, and the symphysial union of the ischia is short.
In all other Mammalia Lepus, Plate 8 [Plate 25], fig. 6), the iliac axis forms as acute, if not a more acute, angle with the sacral axis; the angle between the ilio pectineal axis and the sacral axis more and more approaches a right angle; and that between the sacral axis and the obturator axis becomes more and more acute. The obturator foramen acquires a much larger proportional size. The symphysial union becomes restricted to a greater or less portion of the pubes; or the ventral halves of the ossa innominata may cease to be directly united, even the pubes being far apart in the dry skeleton. The metischial processes are represented by tuberosities, which may extend upwards and unite with anterior caudal vertebræ; and the ilia may remain narrow or become extremely expanded. In all monodelphous Mammalia the marsupial bones disappear.
[347] The distinctive features of the mammalian pelvis have been clearly indicated by Gegenbaur,1 who points out that in mammals, in contradistinction from reptiles, "the longitudinal axis of the ilium gradually acquires an oblique direction, from in front and above, backwards and downwards. The part which represents the crista above thus becomes turned forwards, or more or less outwards, with increase of lateral surface; the acetabular part backwards and downwards; hence the ischium retains its original direction in the produced long axis of the ilium and, at the same time, takes up a position in relation to the vertebral column similar to that which obtains in birds. The conditions of this position are, however, to be sought in factors of a totally different nature in mammals from those which produce it in birds; for in the former, the ischium follows the changed direction of the ilium, whilst in birds, the ilium has nothing to do with the matter, and the ventral elements of the pelvis appear to pass towards the caudal region, independently of the ilium."
On one point, however, I cannot agree with Gegenbaur's conclusions. he is of opinion that the ilium of mammals answers to the post-acetabular part of the ilium of birds, and that "the crista ossis ilii of mammals corresponds with the posterior edge of the postacetabular part of the bird's ilium. Between the two parts, therefore, there is the difference of a rotation through an angle of almost 180°." On the contrary, it appears to me evident that the whole christa ilii in a mammal corresponds with the whole dorsal edge of the ilium in a bird or a reptile, and that the angle through which the iliac axis rotates amounts to not more than 90° (compare Plate 8 [Plate 25], fig. 6, Lepus, with fig. 9, Apteryx). I cannot reconcile the contrary view either with the relations of the ilium to the sacrum, or with the attachment of the muscles.
On comparing the pelvis of Ornithorhynchus with that of a lizard (Plate 8 [Plate 25], fig. 2), or that of a Chelonian, it will be observed that the resemblance between the former and the Sauropsidan pelvis is, in most respects, closer than that which it bears to the higher Mammalian pelvis. In the reptiles both the pubes and the ischia unite in a ventral symphysis; the pubis has a strong pectineal process, which acquires very large dimensions in the Chelonia; the metischial processes are also often very strong. Nevertheless, there is an important difference, for, in all these animals, the iliac axis is either nearly perpendicular to the sacral axis, or slopes from above downwards and forwards; the obturator axis also inclines downwards [348] and forwards. Hence, in most Lacertilia and Chelonia, the pubes slope forwards very obliquely, while the ischia come more and more forwards. In other words, snch modifications of the pelvis as occur in the Lacertilia and the Chelonia are of an opposite kind to those which. take place in Mammalia.
The same thing is true of the Crocodilia (Plate 8 [Plate 25], fig. 3). Here the ilium is much broader than in the lizards and the Chelonia. This broadening is effected by the expansion of the ilium, both in front of and behind the iliac axis, which retains about the same inclination to the sacral axis that it has in lizards. The ischia have but small metischial processes, and their long axes lie further forwards than in most lizards. The obturator axis inclines forwards, and the iliopectineal axis is parallel with the sacral axis, as in lizards. As in Echidna, a space of the inner wall of the acetabulum is fibrous. The lower boundary of this space is constituted by a prolongation of the anterior end of the cotyloid extremity of the ischium. The interval between this and the anterior end of the ilium answers to the cotyloid end of the pubis in a lizard, but it does not ossify. The pubis corresponds exactly in direction with that of a lizard, but its form is very different. At first narrow and rounded, it gradually flattens from above downwards, and, at the same time, widens into a broad trowel-shaped plate of cartilage enclosed in a dense fibrous perichondrium, which lies close beside the middle line in the ventral wall of the abdomen (Plate 8 [Plate 25], fig. 12). Each of these flat cartilages is distinct from its fellow throughout the greater part of its extent but, posteriorly, the two approach, and are united by a broad and strong ligamentous band (Sy,. p. l.). The bony portion of the pubis commences just outside the acetabulum and extends to this band, terminating by a curved edge directed inwards and forwards. It is the osseous portions of the pubes which are commonly described as the entire pubes of the Crocodilia, and much speculative ingenuity has been expended upon the interpretation2 of these apparently anomalous elements of the pelvis, which are readily moveable upon their fibro-cartilagnious connexions with the acetabulum. But in no essential respect do they differ from ordinary pubes. Throughout their whole length they give attachment to a muscle, which answers to the pectineus and short adductors of the [349] thigh, while the aponeurosis which lies between them and the ischia gives origin as usual to the obturator externus; and the obturator nerves pass out close to the cotyloid ends of the pubes (Plate 8 [Plate 25], fig. 12, Ob. n). For the trowel-shaped forward continuations of the pubes on each side of the symphysis, I. will adopt the name of epipubes, proposed by Hoffman for other structures which I believe to be homologous with them. They are firmly connected with the aponeurosis of the external oblique muscle, in which, just in front of their outer edges, lie, on each side, the first set of abdominal false ribs (Plate 8 [Plate 25], fig. 12, r.). In short, in all their most important relations, these appear to me to be structures homologous with the marsupial bones of the Monotremes and Marsupials, which in Thylacinus are represented by mere cartilages. But although homologous, they are very different in detail; and, in all other respects, the Crocodilian pelvis departs even further than that of the Chelonia and Lacertilia from the Mammalian type.
The Pterosauria seem to have possessed epipubes; and in the Dicynodontia there is an approximation to the backward elongation of the subsacral part of the ilium which is characteristic of Mammals; but, in both these groups there appears to have been no obturator fontanelle.
In the ornithoscelidan reptiles and in birds (Plate 8 [Plate 25], figs. 7, 8, and 9), the pelvis, starting from the lacertilian and crocodilian type, undergoes a series of modifications of a new character, the ultimate result of which is a pelvis as much specialised as that of the higher Mammals, but totally different from it in principle. The broadening of the ilium seen in the crocodile increases, so that the antero-posterior length of the bone eventually becomes very great, chiefly in consequence of the elongation of the præaxial region of the ilium. But, with all this, the direction of the iliac axis does not sensibly change, and it remains more or less inclined downwards and forwards (Plate 8 [Plate 25], fig. 9). The inner wall of the acetabulum is largely membranous. The iliopectineal axis becomes slightly inclined to the sacral axis, but never so much even as in Echnida. The main change in the pelvis is, in fact, effected by the extraordinary elongation of the pubes and the ischia, and their rotation backwards and upwards; while, at the same time, the symphysial union of the bones of opposite sides altogether disappears. In Rhea, the ischia unite with some of the post-sacral vertebræ as they do in many Mammalia. The pubis becomes very slender and, as it lies parallel with the ischium, the obturator space is reduced to a [350] mere slit, often bridged over by a process of the ischium.3 The pectineal process is immensely elongated in some Ornithoscelida (as Hulke has shown Iguanodon, and Marsh in Laosaurus (Plate 8 [Plate 25], fig. 8); but, in birds, it is usually short (fig. 9), and may be absent, and no epipubes have been discovered, either in the Ornithoscelida or in birds.4
Thus it appears to be useless to attempt to seek among any known Sauropsida for the kind of pelvis which analogy leads us to expect among those vertebrated animals which immediately preceded the lowest known Mammalia. For, if we prolong the series of observed modifications of the pelvis in this group backwards, the "Promammalia" antecedent to the Monotremes may be expected to have the iliac and obturator axes perpendicular to the sacral axis, and the iliopectineal axis parallel with it; something, in short, between the pelvis of an Ornithorhynchus and that of a land tortoise; and provided, like the former, with large epipubes intermediate in character between those of the lower mammals and those of crocodiles. In fact, we are led to the construction of a common type of pelvis, whence all the modifications known to occur in the Sauropsida and in the Mammalia may have diverged.
It is a well-known peculiarity of the urodele Amphibia, that each os innominatum consists of a continuous cartilage, the ventral half of which is perforated by a foramen for the obturator nerve, but has no large fibrous fontanelle, or obturator foramen, in the ordinary sense of the word. At the junction of the dorsal with the ventral moiety, the acetabulum marks off the iliac portion of the pelvic arch above, from the pubic and ischial regions below; and these are further distinguishable, even apart from their ossifications, by the position of the foramen for the obturator nerve and the origins of the muscles. In full-grown specimens of Salamandra maculosa, the pelvis presents the following characters (Plate 8 [Plate 25 ], figs. 1, 10, and 11):–The iliac axis is slightly inclined forwards, while the iliopectineal axis is practically parallel with the sacral axis. The iliac ossification extends into the acetabulum, and forms a triangular segment of its roof with the apex downwards, exactly as in lizards. The posterior and inferior side of the triangle is separated by a thin band of the primitive cartilage from the upper edge of the similarly triangular [351] cotyloid end of the ischial ossification, the anterior edge of which is vertical, again as in lizards. Between this edge and the anterior and inferior edge of the iliac ossification there is a cartilaginous interspace, as in crocodiles, which represents the cotyloid end of the pubis (Plate 8 [Plate 25], fig. 1). This cartilaginous part of the pubis gives rise to a pectineal process (p. p.), which has the same position as in birds and in Ornithorhynchus. In the floor of the acetabulum, the pubic ossification (fig. 1, Pb.) makes its appearance as a very thin lamina, which extends, underneath the pectineal process, inwards; and gradually surrounds the whole of the thickened transverse ridge of cartilage which corresponds with the pubis. The pubis is thus represented by an axis of cartilage surrounded by bone, and the thick inner extremities of the two pubes are largely united by fibrous tissue (fig. 10, Sy. p.). The ischia are relatively large, and are united, partly by cartilage and partly by ligament, in a long symphysis (fig. 10, Sy. I). Their posterior and external angles are produced into short metischial processes. In one specimen, I observed a distinct sutural line (Plate 8 [Plate 25], fig. 11 s.), between the anterior curved edge of the right ischium and the corresponding pubis, while no such suture could be traced upon the other side.
The pelvic arch of Salamandra, therefore, contains all the elements which are found in the higher Vertebrata, but the obturator fontanelle is wanting.
Proceeding from the symphysis pubis, with which it is connected by ligament, is the "ypsiloid" cartilage or epipubis (figs. 1, 11, Ep. p.), which was called by the accurate and acute Dugès the "marsupial cartilage." This indication of the homology of the part has been adopted by Cuvier and others, but I do not know that it has been generally accepted. I believe, however, that the identification is perfectly just.
The ypsiloid cartilage proceeds forwards in the middle line, as a stem (Ep. p.), of variable length, and then divides into two branches (Ep. p1., Ep2.), which diverge at right angles to one another, and terminate by rounded extremities. The pedicle of the ypsiloid cartilage is broad and triangular in section, the apex of the triangle being dorsal.
The manner in which the abdominal muscles are connected with this cartilage is very instructive (Plate 8 [Plate 25], figs. 13. 15). The anterior pectinations of the external oblique muscle (0, e.) are inserted into a thin but dense fascia, which is hardly separable from the superjacent dermis, and which extends across the middle line to the muscle of the other side. The most [352] posterior bundles of the external oblique, however, are inserted by a rounded tendon into the pectineal process (Plate 8 [Plate 251, fig. 15, p. p.), which therefore answers to the spine or tuberosity of the pubis in Mammalia. Internal to this, the fascia arches over a small space (i. r.) (which corresponds with the inguinal ring), and is then inserted into the whole length of the pedicle and into the posterior half of each ramus of the ypsiloid cartilage. Beyond this point the rami are free externally, though they lie close against the fascia. On their inner sides, however, they are connected together and with the fascia of the external oblique, in the middle line, by a delicate sheet of fibrous tissue (figs. 13 and 15, f). On comparing this disposition of the tendon of the external oblique muscle with that which obtains in Ornithorhynchus (Plate 8 [Plate 25], fig 14) the correspondence is obvious. For, in the latter, the posterior fibres of that muscle are inserted directly into the spine of the pubis (t. p.). Between these and the outer edge of the marsupial bone there is fibrous interspace, corresponding with the inguinal ring (i. r.) ; while the more anterior fibres of the external oblique are inserted into the apex of the bone, which is, as it were, imbedded in the fascia. If the pedicle of the ypsiloid cartilage were reduced, the rami at the same time widening behind, until their outer angles reached the pectineal processes, and their free apices shortening the ypsiloid cartilage of the salamander would be converted into two cartilages having exactly the same relation to the tendon of the external oblique that the marsupial bones have in the Ornithorhynchus.
In the Monotremes (Plate 8 [Plate 251, fig. 14, Py..), there are two very large pyramidales muscles, which spring from the whole inner margins of the marsupial bones; their posterior and middle fibres run to the middle line, but the anterior ones constitute a longitudinal band, which extends forwards, and is inserted along with the rectus (R.), of which, indeed, it looks like a part. In the salamander, muscular fibres similarly take their origin from the inner edges of the rami of the epipubis; and the most anterior of these fibres pass forwards, and become more or less confounded with the inner edges of the recti (fig. 13, Py.). But the region which answers to the linea alba (l. a.) is very broad, whence the pyramidales are separated by a wide interval.
Thus far, the muscles which are connected with the epipubis are strictly comparable with those which are attached to the marsupial bones. But, in Salamandra, there is a muscle (Plate 8 [Plate 25], fig. 13, A.), of which I have been able to find no representative in the [353] Monotremes I have dissected. This is a thin band of longitudinal fibres, which spring partly from the pubis, close to the symphysis, and partly from the outer edge of the pedicle of the epipubis, and are inserted into the outer edge of the ramus (Plate 8 [Plate 25], fig. 13, A.). External to this, a broad flat band of muscular fibres (fig. 13, B.) takes its origin from the pubis and its pectineal process, and runs forwards to be inserted into the modified branchial arches and the tongue. These are the hebosteoglossi of von Siebold ("Observationes quædam de Salamandris et Tritonibus"). Superficial to this muscle, is the proper rectus (R.) itself, marked by its tendinous intersections; while, on the dorsal or deep side of both, there is a curious fanshaped muscle (C.), which takes its origin from the pectineal process, by a narrow tendon, and spreads out to be inserted into the outer face of the pedicle and the outer edge of the ramus of its side. The ventral face of the pedicle and the posterior halves of the rami are closely adherent to the fascia of the external oblique in the middle line, and lie between the inner edges of the two recti. Finally, the fibres of the transversus (fig. 13, Trs .) are inserted into the outer edges of the rami, and the fascia which unites these appears to belong to the transversus.
On comparing the recti and the muscles A, B, C, with those of Ornithorhynchus and Echidna, a great apparent difference manifests itself. For the very thin recti of the Monotremes take their origin from the pubis along a line which extends from the tubercle to close to the symphysis, and pass forwards, dorsad of the marsupial bones and the pyramidales, which thus lie altogether in front of them and are, by them, largely separated from the fascia transversalis. Nevertheless, it will be observed that the origin of the muscle B nearly corresponds with that of the rectus in Ornithorhynchus; and I am disposed to think that, in this animal, the rectus, at least in its posterior moiety, is represented by the homologue of this muscle, which has extended laterally over the dorsal face of the enormously enlarged homologues of the rami of the ypsiloid cartilage.
However this may be, it must be recollected that it is only the extreme ends of the rami which lie dorsad of the recti, and that, in the rest of its extent, the epipubis of Salamandra is firmly fixed to the fascia of the external oblique, which forms the front wall of the sheath of the rectus. The homology of the epipubis with the marsupial bones is determined by the essential identity of the relations of the two to the tendons of the external oblique muscles.
It seems to me that, in such a pelvis as that of Salamandra, we [354] have an adequate representation of the type from which all the different modifications which we find in the higher Vertebrata may have taken their origin.
In the lizards and the Chelonia the iliac and obturator axes have inclined forwards, and the epipubes have been reduced to such rudiments as have been described in chameleons and in some tortoises.5
In the crocodiles, with the same general pelvic characters, the cotyloid end of the pubis retains its imperfectly ossified condition, while the epipubes represent the vastly enlarged rami of the salamandrine epipubis.
In the Ornithoscelida and in birds, the ilia elongate, but it is the modification of the pubes and ischia which is the most characteristic feature of the pelvis, and the epipubis vanishes.
In the Pterosauria and in the Dicynodonts, the salamandrine non-development of an obturator fontanelle persists;; and in the former, the sessile rami of the epipubis appear to be represented by the so-called marsupial bones.
Unless the like should prove to be the case in the Dicynodonts, it is in the Mammalia alone that the subsacral portion of the ilium elongates backwards, carrying with it the pubis and the ischium, between which a large rounded obturator fontanelle is developed.
These facts appear to me to point to the conclusion that the Mammalia have been connected with the Amphibia by some unknown "pro-mammalian" group, and not by any of the known forms of Sauropsida, and there is other evidence which tends in the same direction.
Thus, the Amphibia are the only air-breathing Vertebrata, which, like Mammals, have a dicondylian skull. It is only in them that the articular element of the mandibular arch remains cartilaginous; while the quadrate ossification is small, and the squamosal extends down over it to the osseous elements of the mandible; thus affording an easy transition to the mammalian condition of these parts.
The pectoral arch of the Monotremes is as much amphibian as it is sauropsidan; the carpus and the tarsus of all Sauropsida, except the Chelonia, are modified away from the Urodele type, while those of the Mammal are directly reducible to it; and it is perhaps worth notice, that the calcar of the frogs is, in some respects, comparable with the spur of the Monotremes.
[355] Finally, the fact that, in all Sauropsida, it is a right aortic arch which is the main conduit of arterial blood leaving the heart, while in Mammals, it is a left aortic arch which performs this office, is a great stumbling-block in the way of the derivation of the Mammalia from any of the Sauropsida. But, if we suppose the earliest forms of both the Mammalia and the Sauropsida to have had a common Amphibian origin, there is no difficulty in the supposition that, from the first, it was a left aortic arch in the one series, and the corresponding right aortic arch in the other, which became the predominant feeder of the arterial system.
The discovery of the intermediate links between Reptilia and Aves, among extinct forms of life, gives every ground for hoping that, before long, the transition between the lowest Mammalia at present known and the simpler Vertebrata may be similarly traced. The preceding remarks are intended to direct attention to the indications of the characters of these promammalian Vertebrata, which the evidence at present forthcoming seems to me to suggest.
In the relatively large size of the brain, and in the absence of teeth, the only existing representatives of the Ornithodelphia present characters which suggest that they are much modified members of the group. On comparing the brain of Echidna, for example, with that of many Marsupialia and Inisectivora, its relative magnitude is remarkable: and, in view of the evidence which is now accumulating, that the brain increases in size in the later members of the same series of Mammalia, one may surmise that Echnida is the last term of a series of smaller-brained Ornithodelphia. Among the Higher Vertebrata, I think that there is strong reason to believe that edentulous animals are always modifications of toothed forms.
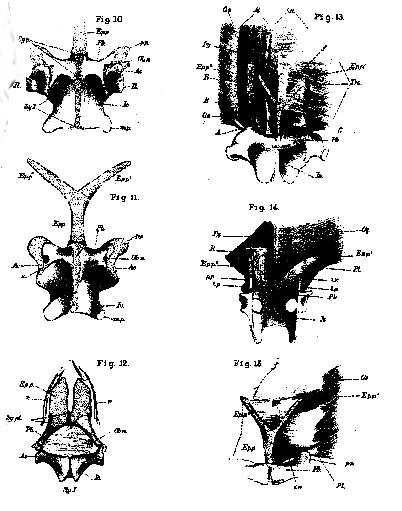
EXPLANATION OF PLATE 8 [PLATE 25].
Figs. 1 to 9. The left half of the pubic arch in Salamandra (fig. 1), Iguana (fig. 2), Crocodilus (fig. 3), Ornithorhynchus (fig. 4), Echidna (fig. 5), Lepus (fig. 6), Compsognathus (fig. 7), Laosaurus (fig. 8), and Apteryx (fig. 9). The letters have the same signification throughout. Il. ilium, Pb. pubis, Is. ischium. Ep.p. epipubis, S. a. sacral axis, Ip. a ilio-pectoral axis, Ob. a. obturator axis, Il. a. ilia axis, Sy. p., Sy. I., indicate the extent to which the pubes and the ischia unite respectively in their ventral symphyses; p. p. pectineal process, t. p. tuberosity or spine of the pubis, . m. p. metischial process or tuberosity of the ischium, Cl. os cloacæ in Iguana.
Figs. 10, 11. Dorsal and ventral views of the ventral half of the pelvis of Salamandra maculosa (x4). The letters as before, except Epp.1 Epp.2, right and left rami of the epipubis or ypsiloid cartilage, Ob. n. foramen of the obturator nerve, s. trace of a suture between the ischium and the pubis, on the right side, Ac. acetabulum.
[356] Fig. 12. Ventral aspect of the pelvis of a small Crocodilus acutus and the hindermost abdominal false ribs, of the natural size. The obturator nerve (0b. n.) perforates the aponeurosis, which fills up the rhomboidal space between the ischia and the pubes, on the inner side of the pubis. Sy. p. l. the ligamentous union of the pubes.
Fig. 12. Dorsal aspect of the epipubis and the muscles connected with it in Salamandra maculosa. On the left side, the transversalis and the muscle C. are removed.
Fig. 13. The same, all the muscles but the external oblique being removed..
Fig. 14. The epipubis and the adjacent muscles in Ornithorhynchus.
R. rectus abdomnus, Py. pyramidalis, O. e. obliquus externus, Trs. transversus, l. a. linea alba, f. fascia, extending between the two rami of the epipubis in Salamandra, P. l. the representative of Poupart's ligament, i. r. inguinal ring, A. B. C. muscles of Salamandra described in the text. A, and C. together appear to be the pubio-marsupial of Dugès; B is the hebosteoglossus of von Siebold.
1 "Beiträge zur Kenntniss des Beckens der Vögel," "Jenaische Zeitschrift," vi. 2 For the latest of these interpretations see Hoffman's excellent memoir, "Beiträge zur Kenntniss des Beckens der Amphibien und Reptilien." "Nied. Archiv für Zoologie," 1876. I cannot but think that had Professor Hoffman studied the crocodile's pelvis in fresh or spirit specimens, he would not have ptit forsvard the hypothesis that the pubes of the crocodiles are "epipubes." Rathke's account of the development of the crocodile's pelvis affords conclusive evidence respecting the homologies of the pubes in these animals. 3 I was, at one time, inclined to think that this represented the union of the pubes and ischia of the same side in ordinary Sauropsida, and that the rest of the ischium represented an unusually elongated metischial process; bnt the study of the development of the pelvis in the chick has convincecl me that this is not the case, 4 See, however, the observations of Mr. Garrod on a "marsupial bone" in ostriches. "Proceedings of the Zoological Society," 1872. 5 Hoffman, "Beiträge zur Kenntniss des Beckens der Amphibien und Reptilian," Neid. Archiv für Zoologie," Bd. 3, p. 143, 1876.
|
THE
HUXLEY
FILE

|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
{kind=link}